

HIF1α-regulated glycolysis promotes activation-induced cell death and IFN-γ induction in hypoxic T cells
- PMID: 39477954
- PMCID: PMC11526104
- DOI: 10.1038/s41467-024-53593-8
HIF1α-regulated glycolysis promotes activation-induced cell death and IFN-γ induction in hypoxic T cells
Abstract
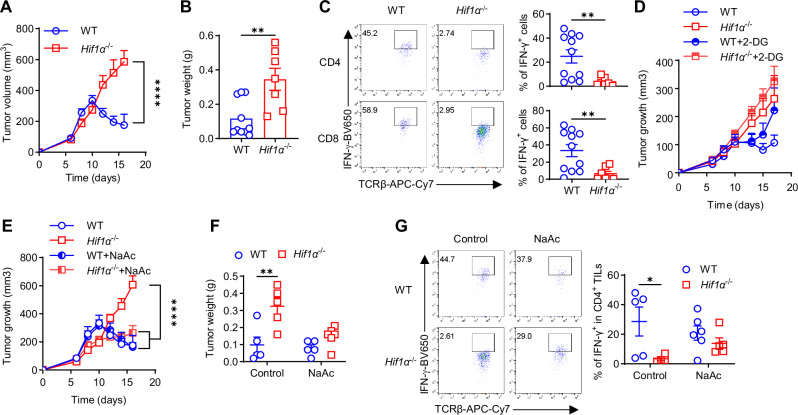
Hypoxia is a common feature in various pathophysiological contexts, including tumor microenvironment, and IFN-γ is instrumental for anti-tumor immunity. HIF1α has long been known as a primary regulator of cellular adaptive responses to hypoxia, but its role in IFN-γ induction in hypoxic T cells is unknown. Here, we show that the HIF1α-glycolysis axis controls IFN-γ induction in both human and mouse T cells, activated under hypoxia. Specific deletion of HIF1α in T cells (Hif1α-/-) and glycolytic inhibition suppresses IFN-γ induction. Conversely, HIF1α stabilization by hypoxia and VHL deletion in T cells (Vhl-/-) increases IFN-γ production. Hypoxic Hif1α-/- T cells are less able to kill tumor cells in vitro, and tumor-bearing Hif1α-/- mice are not responsive to immune checkpoint blockade (ICB) therapy in vivo. Mechanistically, loss of HIF1α greatly diminishes glycolytic activity in hypoxic T cells, resulting in depleted intracellular acetyl-CoA and attenuated activation-induced cell death (AICD). Restoration of intracellular acetyl-CoA by acetate supplementation re-engages AICD, rescuing IFN-γ production in hypoxic Hif1α-/- T cells and re-sensitizing Hif1α-/- tumor-bearing mice to ICB. In summary, we identify HIF1α-regulated glycolysis as a key metabolic control of IFN-γ production in hypoxic T cells and ICB response.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Update of
-
HIF1α-glycolysis engages activation-induced cell death to drive IFN-γ induction in hypoxic T cells.Res Sq [Preprint]. 2024 Jan 12:rs.3.rs-3830704. doi: 10.21203/rs.3.rs-3830704/v1. Res Sq. 2024. Update in: Nat Commun. 2024 Oct 30;15(1):9394. doi: 10.1038/s41467-024-53593-8. PMID: 38260594 Free PMC article. Updated. Preprint.
References
-
- Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M. & Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol.155, 1151–1164 (1995). - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- 1R21CA259721-01A1/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- CRI4342/Cancer Research Institute (CRI)
- 1R01CA279849-01A1/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- O'Neal Invests Pre-R01 Award/UAB | School of Medicine, University of Alabama at Birmingham (UAB School of Medicine)
- R01 CA279849/CA/NCI NIH HHS/United States
- R21 CA259721/CA/NCI NIH HHS/United States
- R21 CA230475/CA/NCI NIH HHS/United States
- 1R21CA230475-01A1/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- V2018-023/V Foundation for Cancer Research (V Foundation)
- ME210108/U.S. Department of Defense (United States Department of Defense)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
