

RNA-binding protein YBX3 promotes PPARγ-SLC3A2 mediated BCAA metabolism fueling brown adipogenesis and thermogenesis
- PMID: 39481849
- PMCID: PMC11570976
- DOI: 10.1016/j.molmet.2024.102053
RNA-binding protein YBX3 promotes PPARγ-SLC3A2 mediated BCAA metabolism fueling brown adipogenesis and thermogenesis
Abstract
Objective: Activating brown adipose tissue (BAT) thermogenesis is a promising approach to combat obesity and metabolic disorders. The post-transcriptional regulation of BAT thermogenesis mediated by RNA-binding proteins (RBPs) is still not fully understood. This study explores the physiological role of novel RBPs in BAT differentiation and thermogenesis.
Methods: We used multiple public datasets to screen out novel RBPs responsible for BAT differentiation and thermogenesis. In vitro loss- and gain-of-function experiments were performed in both C3H10T1/2 preadipocytes and mature brown adipocytes to determine the role of Y-box binding protein 3 (YBX3) in brown adipocyte differentiation and thermogenesis. Adeno-associated virus (AAV)-mediated BAT-specific knockdown or overexpression of Ybx3 was applied to investigate the function of YBX3 in vivo.
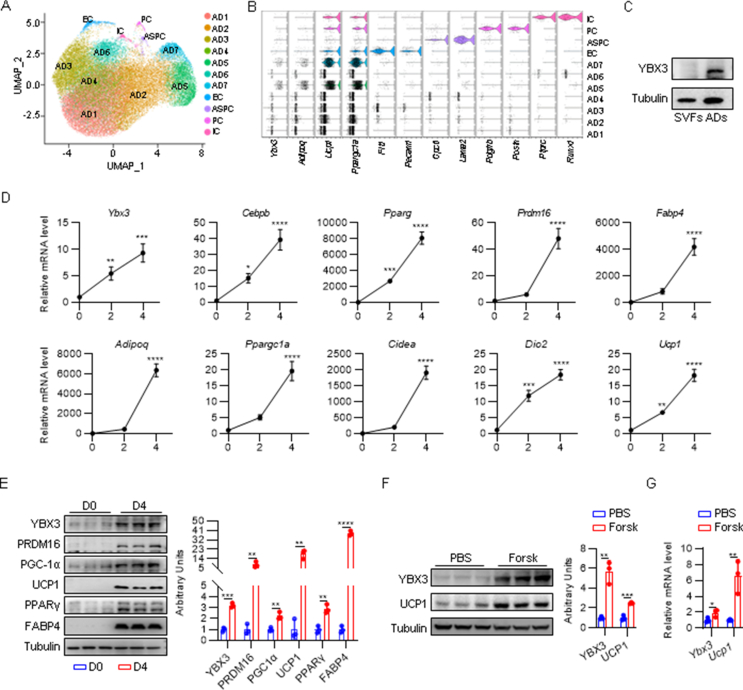
Results: YBX3 is a brown adipocyte-enriched RBP induced by cold stimulation and β-adrenergic signaling. Both in vitro loss- and gain-of-function experiments demonstrate that YBX3 is essential for brown adipocyte differentiation and thermogenesis. BAT-specific loss of Ybx3 dampens thermogenesis and exacerbates diet-induced obesity in mice, while overexpression of Ybx3 promotes thermogenesis and confers protection against diet-induced metabolic dysfunction. Transcriptome analysis and mitochondrial stress test indicate that Ybx3 deficiency compromises the mitochondrial oxidative phosphorylation, leading to thermogenic failure. Mechanistically, YBX3 stabilizes the mRNA of Slc3a2 and Pparg, which facilitates branched-chain amino acid (BCAA) influx and catabolism and fuels brown adipocyte differentiation and thermogenesis.
Conclusions: YBX3 facilitates BAT fueling BCAA to boost thermogenesis and energy expenditure, which protects against obesity and metabolic dysfunction. Thus, YBX3 could be a promising therapeutic target for obesity.
Keywords: Branched-chain amino acid; Brown adipose tissue; Obesity; Thermogenesis; YBX3.
Copyright © 2024 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures












References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
