

GLP-1 and its derived peptides mediate pain relief through direct TRPV1 inhibition without affecting thermoregulation
- PMID: 39482537
- PMCID: PMC11612315
- DOI: 10.1038/s12276-024-01342-8
GLP-1 and its derived peptides mediate pain relief through direct TRPV1 inhibition without affecting thermoregulation
Abstract
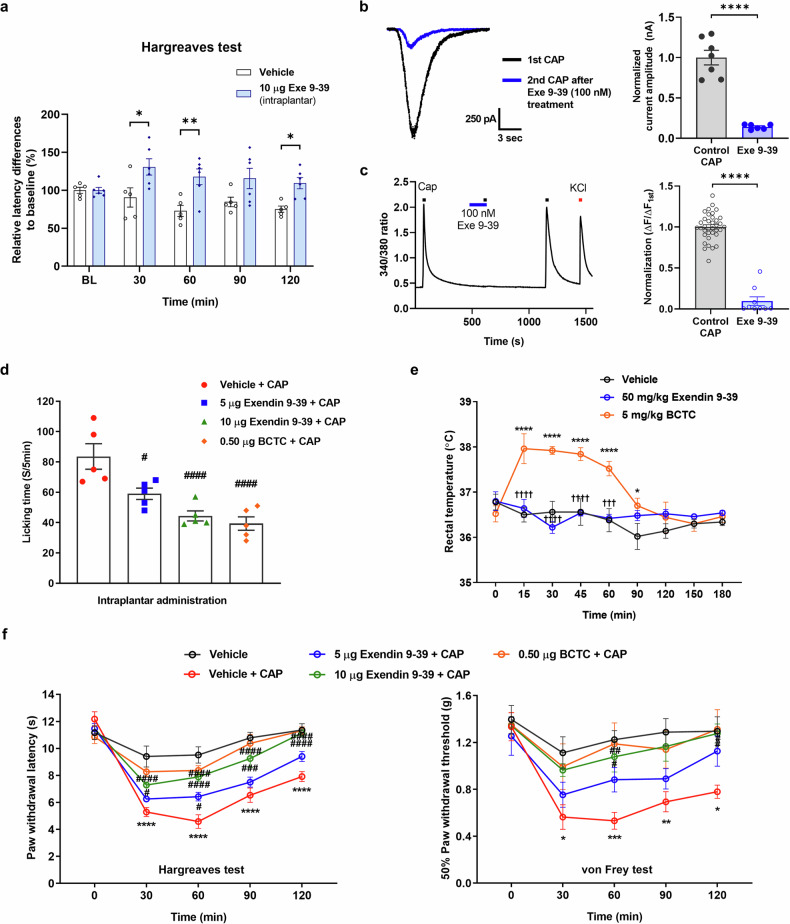
Hormonal regulation during food ingestion and its association with pain prompted the investigation of the impact of glucagon-like peptide-1 (GLP-1) on transient receptor potential vanilloid 1 (TRPV1). Both endogenous and synthetic GLP-1, as well as a GLP-1R antagonist, exendin 9-39, reduced heat sensitivity in naïve mice. GLP-1-derived peptides (liraglutide, exendin-4, and exendin 9-39) effectively inhibited capsaicin (CAP)-induced currents and calcium responses in cultured sensory neurons and TRPV1-expressing cell lines. Notably, exendin 9-39 alleviated CAP-induced acute pain, as well as chronic pain induced by complete Freund's adjuvant (CFA) and spared nerve injury (SNI), in mice without causing hyperthermia associated with other TRPV1 inhibitors. Electrophysiological analyses revealed that exendin 9-39 binds to the extracellular side of TRPV1, functioning as a noncompetitive inhibitor of CAP. Exendin 9-39 did not affect proton-induced TRPV1 activation, suggesting its selective antagonism. Among the exendin 9-39 fragments, exendin 20-29 specifically binds to TRPV1, alleviating pain in both acute and chronic pain models without interfering with GLP-1R function. Our study revealed a novel role for GLP-1 and its derivatives in pain relief, suggesting exendin 20-29 as a promising therapeutic candidate.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures







Update of
-
GLP-1 and Its Derived Peptides Mediate Pain Relief Through Direct TRPV1 Inhibition Without Affecting Thermoregulation.Res Sq [Preprint]. 2024 May 13:rs.3.rs-4233732. doi: 10.21203/rs.3.rs-4233732/v1. Res Sq. 2024. Update in: Exp Mol Med. 2024 Nov;56(11):2449-2464. doi: 10.1038/s12276-024-01342-8. PMID: 38798444 Free PMC article. Updated. Preprint.
References
-
- Meleger, A. L., Froude, C. K. & Walker, J. 3rd. Nutrition and eating behavior in patients with chronic pain receiving long-term opioid therapy. PM R.6, 7–12.e11 (2014). - PubMed
-
- Amy Janke, E. & Kozak, A. T. “The more pain I have, the more I want to eat”: obesity in the context of chronic pain. Obesity20, 2027–2034 (2012). - PubMed
-
- Casey, K. L. & Morrow, T. J. Nocifensive responses to cutaneous thermal stimuli in the cat: stimulus-response profiles, latencies, and afferent activity. J. Neurophysiol.50, 1497–1515 (1983). - PubMed
-
- Wylie, L. M. & Gentle, M. J. Feeding-induced tonic pain suppression in the chicken: reversal by naloxone. Physiol. Behav.64, 27–30 (1998). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
