

Genome-wide identification of rice CXE gene family and mining of alleles for potential application in rice improvement
- PMID: 39483679
- PMCID: PMC11524881
- DOI: 10.3389/fpls.2024.1435420
Genome-wide identification of rice CXE gene family and mining of alleles for potential application in rice improvement
Abstract
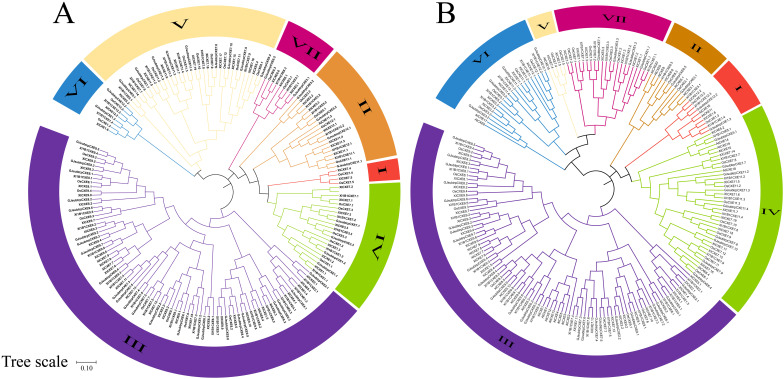
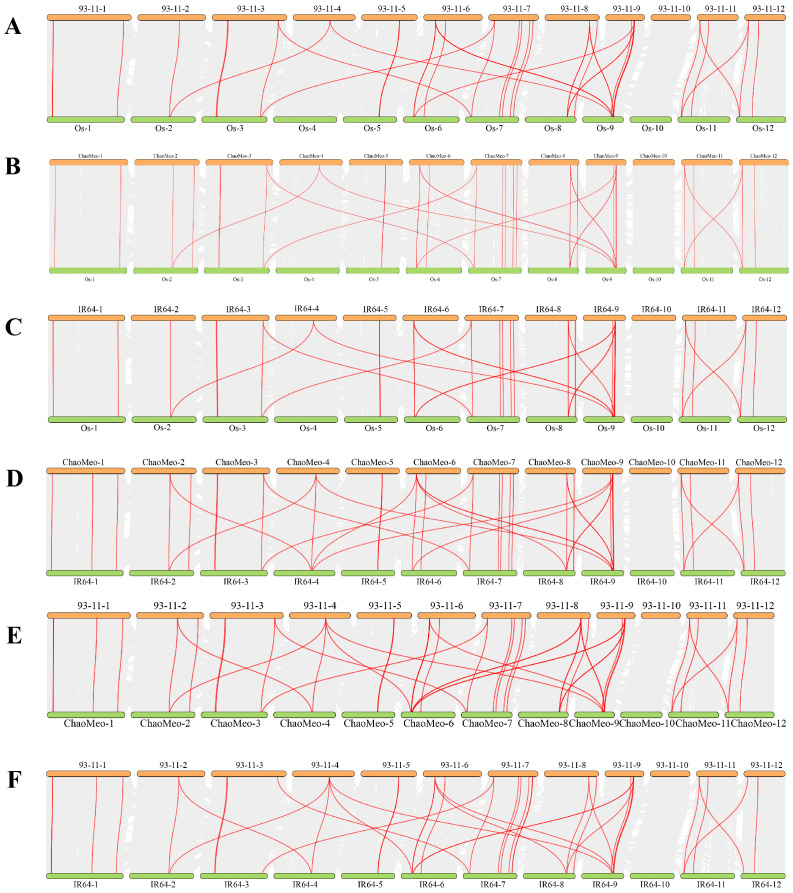
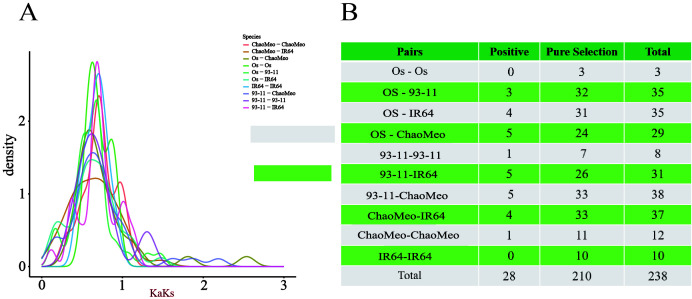
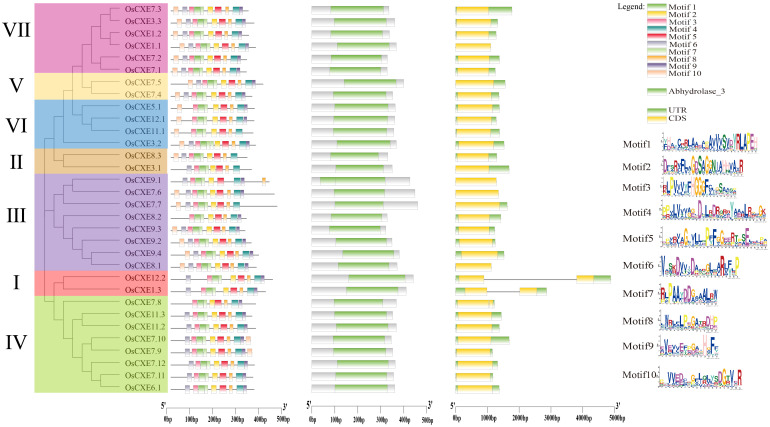
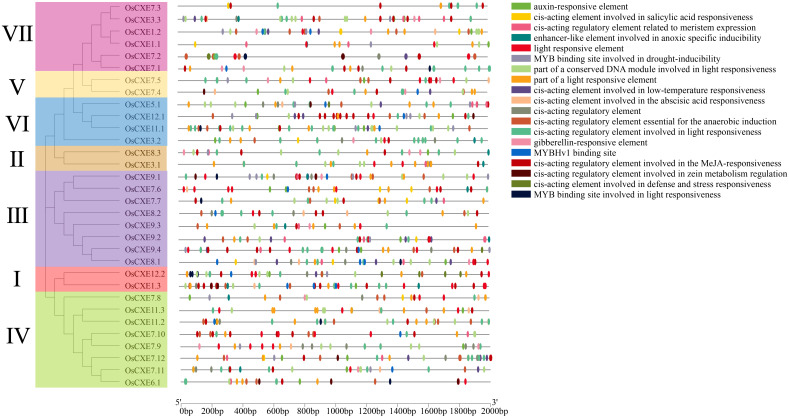
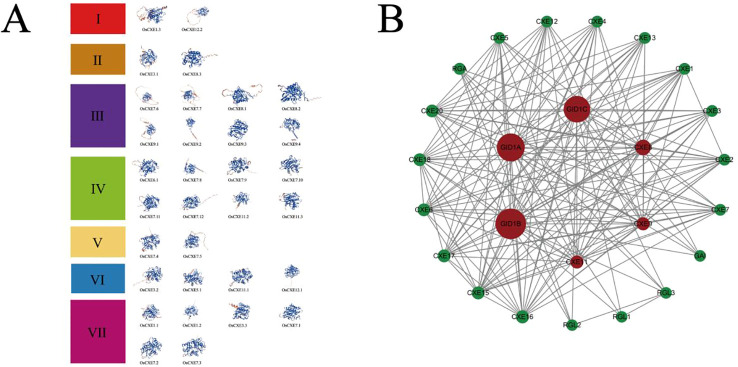
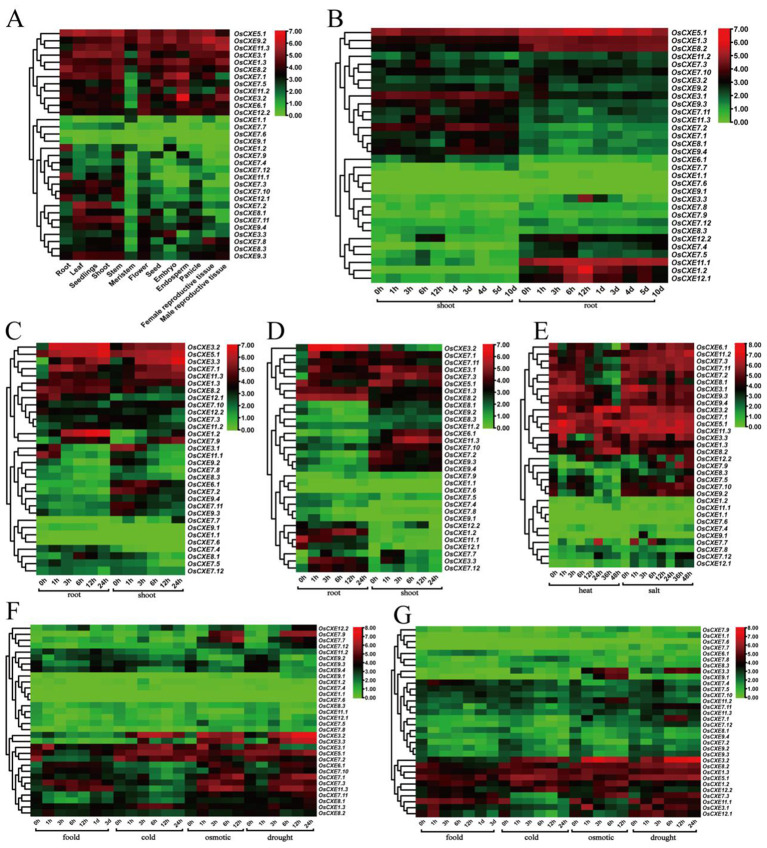
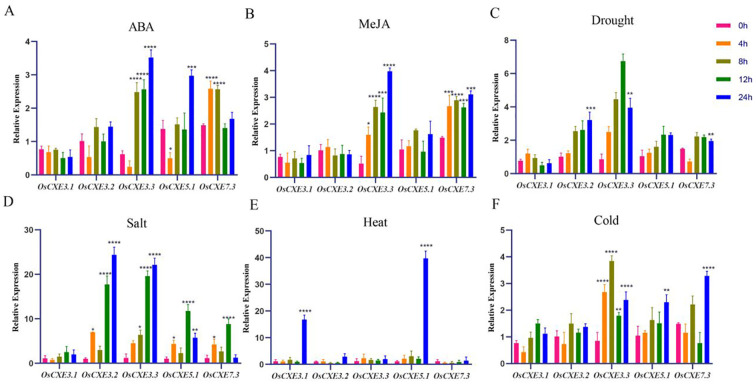
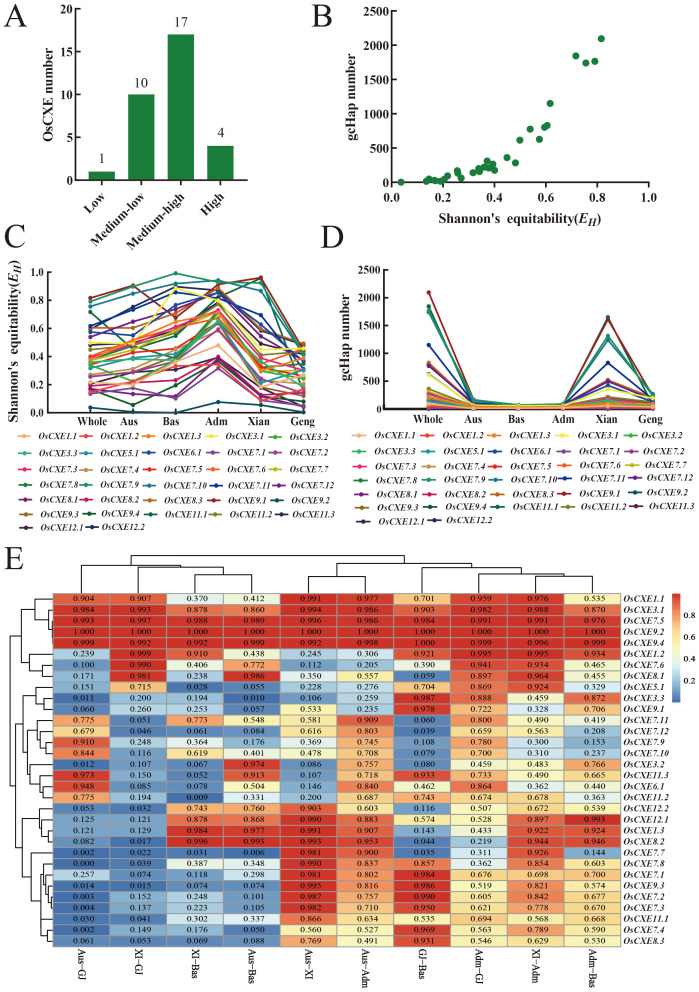
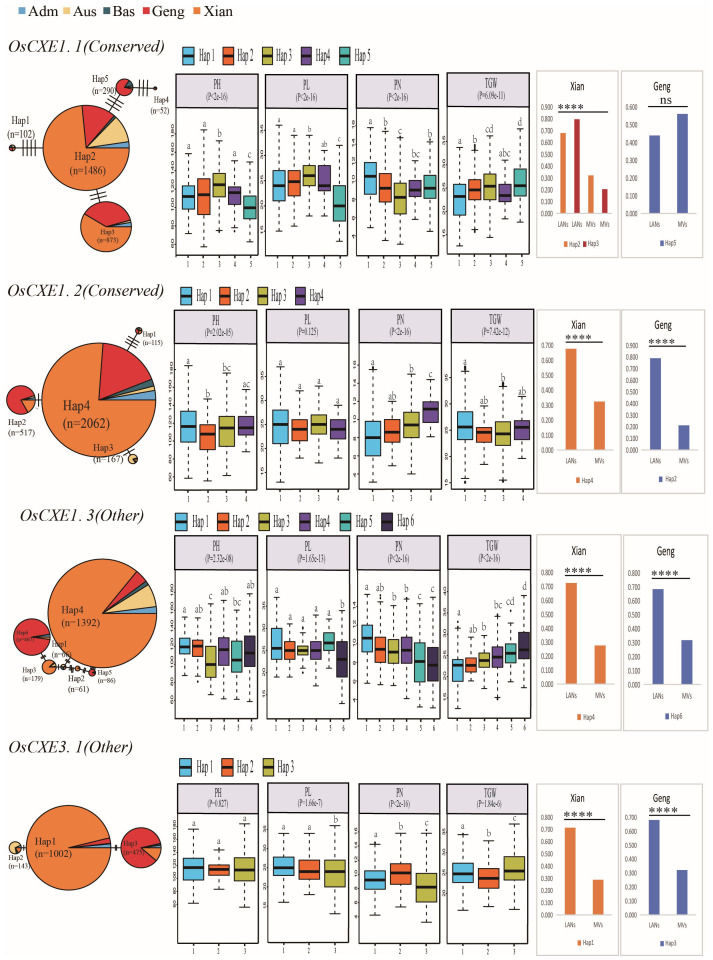
Carboxylesterases (CXE, EC 3.1.1.1), a class of hydrolases with an α/β folding domain, play important roles in plant growth and development and stress response. Here, we identified 32, 63, 41, and 45 CXE genes in Oryza sativa Japonica (Nipponbare), Oryza sativa Indica (93-11), Oryza sativa Indica (Xian-1B1 var.IR64), and Oryza sativa Japonica (Geng-sbtrp var.ChaoMeo), respectively. Then, we analyzed the chromosomal location, physical and chemical properties, subcellular localization, collinearity, and selection pressure of CXE genes in four rice varieties. We also analyzed the functional interaction network, cis-regulatory elements, evolutionary relationship, and protein tertiary structure, and performed gene expression profiling and qPCR verification under abiotic stress, as well as diversity analysis of 3010 gene-CDS-haplotype (gcHap) rice samples, aiming to understand the potential function of the 32 OsCXE genes. Our results indicated that fragment replication is the main reason for amplification of the CXE gene family in rice, and the gene family has undergone strong purification selection. OsCXE3.1, OsCXE3.2, OsCXE3.3, OsCXE5.1, and OsCXE7.3 may be used to improve the tolerance of rice to abiotic stress. OsCXE play important roles in rice population differentiation and improvement, and the major gcHaps at most OsCXE locus are significantly associated with yield traits. Therefore, natural variations of most OsCXE locus have great potential value for improvement of rice productivity.
Keywords: biological stress; carboxylesterase; gene-CDS-haplotype (gcHap) diversity; rice; yield traits.
Copyright © 2024 Zhang, Wang, Dou, Meng, Tang, Lv, Wang, Wang, Li, Bao, Zhang, Huang and Shi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
LinkOut - more resources
Full Text Sources
Research Materials