

This is a preprint.
NINJ1 regulates plasma membrane fragility under mechanical tension
- PMID: 39483869
- PMCID: PMC11527205
- DOI: 10.21203/rs.3.rs-5237916/v1
NINJ1 regulates plasma membrane fragility under mechanical tension
Update in
-
NINJ1 regulates plasma membrane fragility under mechanical strain.Nature. 2025 Aug;644(8078):1088-1096. doi: 10.1038/s41586-025-09222-5. Epub 2025 Jun 9. Nature. 2025. PMID: 40490006 Free PMC article.
Abstract
Plasma membrane integrity is vital not only for cell survival but also nearly all aspects of cell functioning1. Mechanical stress can cause plasma membrane damage2, but it is not known whether there are large molecules (proteins) that control plasma membrane integrity. Here we constructed a 384-well cellular stretch system that delivers precise, reproducible mechanical strain to adherent cells. Using the system, we screened 10,843 siRNAs targeting 2,726 multi-pass transmembrane proteins for stretch-induced membrane permeability changes. The screen identified NINJ1, a protein recently proposed to regulate pyroptosis and other lytic cell death3,4, as the top hit. We demonstrate that NINJ1 is a critical regulator for mechanical force-induced plasma membrane rupture (PMR), without the need of stimulating any cell death programs. Low NINJ1 expression renders the membrane more resistant to stretching, while high expression of NINJ1 lowers the threshold of PMR under mechanical strain. NINJ1 level on the plasma membrane is inversely correlated to tension required to rupture the membrane. In the pyroptosis context, NINJ1 on its own is not sufficient to fully rupture the membrane, and additional mechanical stress is required for full PMR. Our work establishes that NINJ1 functions as a bona fide determinant of membrane biomechanical properties. Our study also suggests that PMR across tissues of distinct mechanical environments is subjected to fine tuning by differences in NINJ1 expression and external mechanical forces.
Conflict of interest statement
Competing Interests J.B., C.P. and S.H are employees of Novartis Biomedical Research. F.S. is the scientific founder and chair of the scientific advisory board of Pyrotech Therapeutics. Other authors declare no competing final interest.
Figures







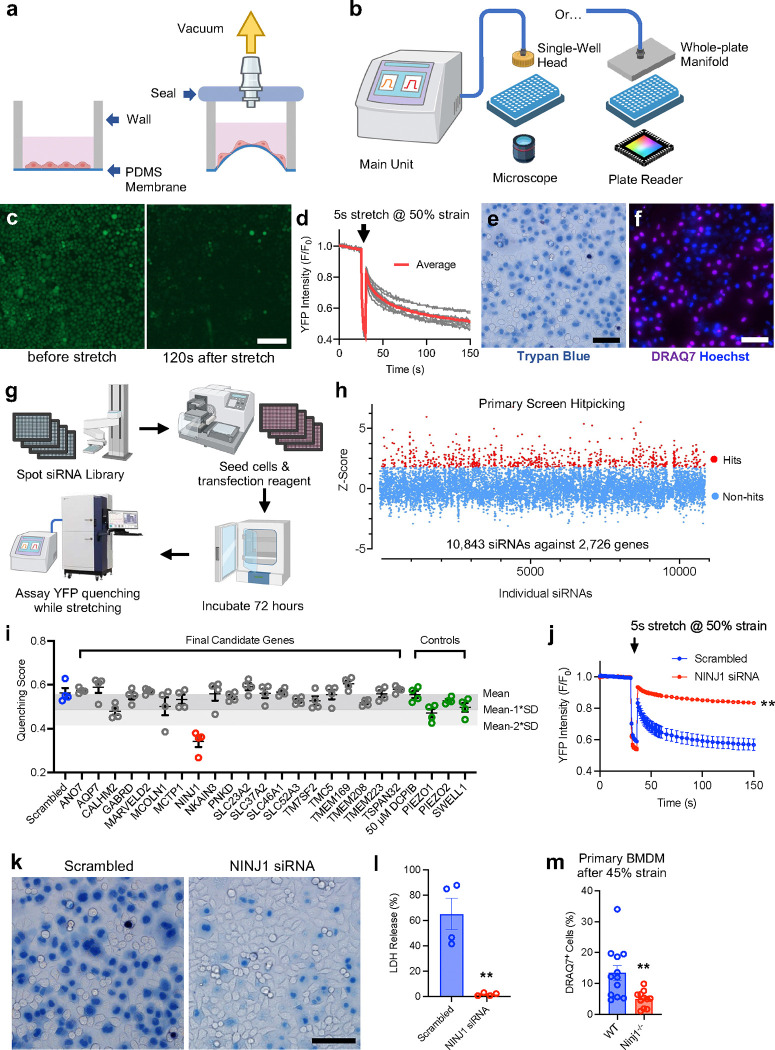



References
-
- Alberts B., Johnson A., Lewis J. in Molecular Biology of The Cell (Garland Science, 2002).
-
- Kayagaki N. et al. NINJ1 mediates plasma membrane rupture during lytic cell death. Nature 591, 131–136 (2021). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
