

This is a preprint.
Decoding the transcriptomic signatures of psychological trauma in human cortex and amygdala
- PMID: 39484441
- PMCID: PMC11526900
- DOI: 10.1101/2024.10.23.619681
Decoding the transcriptomic signatures of psychological trauma in human cortex and amygdala
Abstract
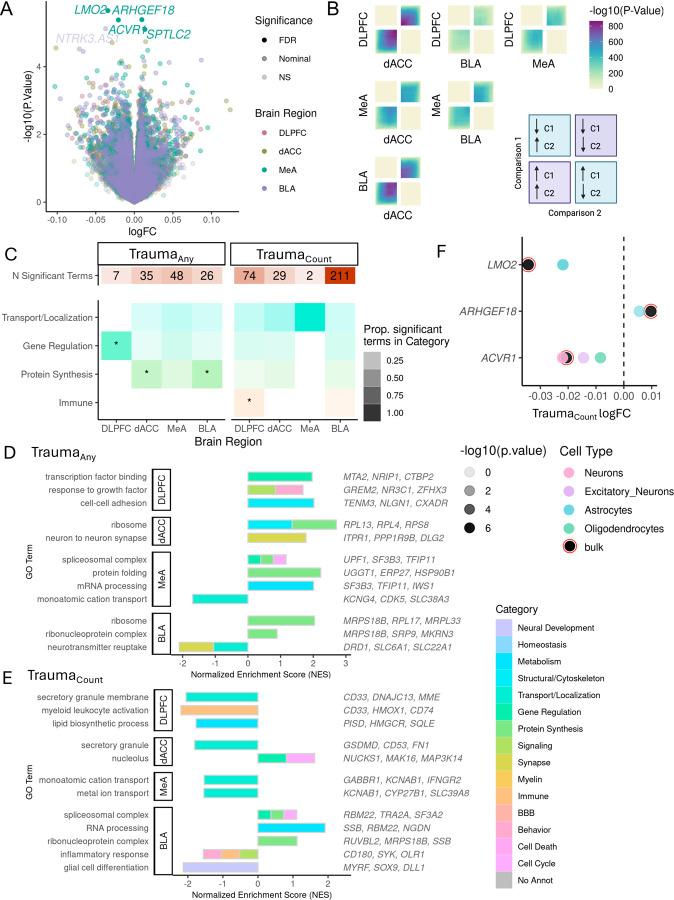
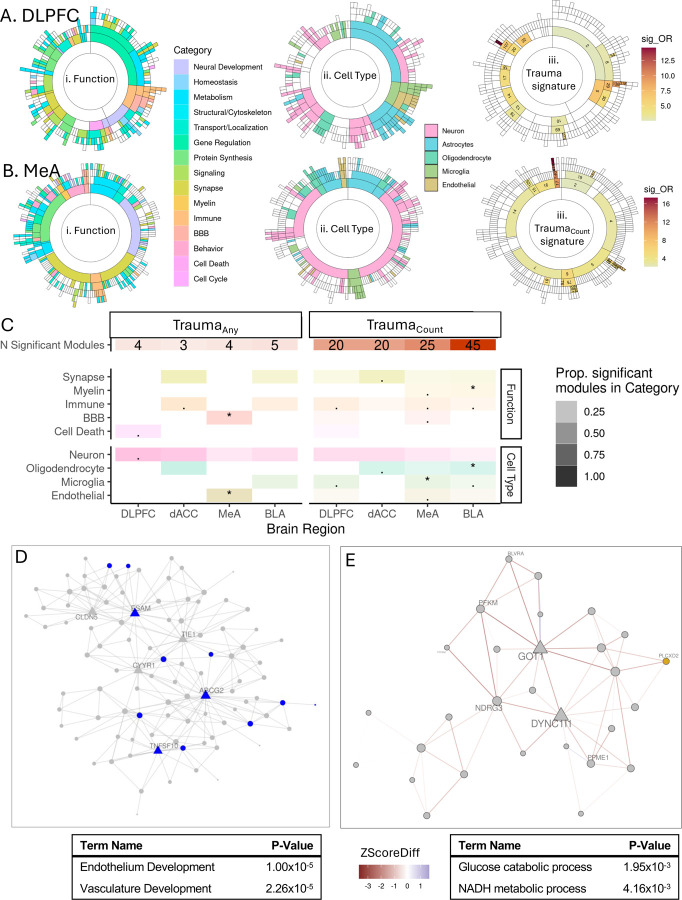
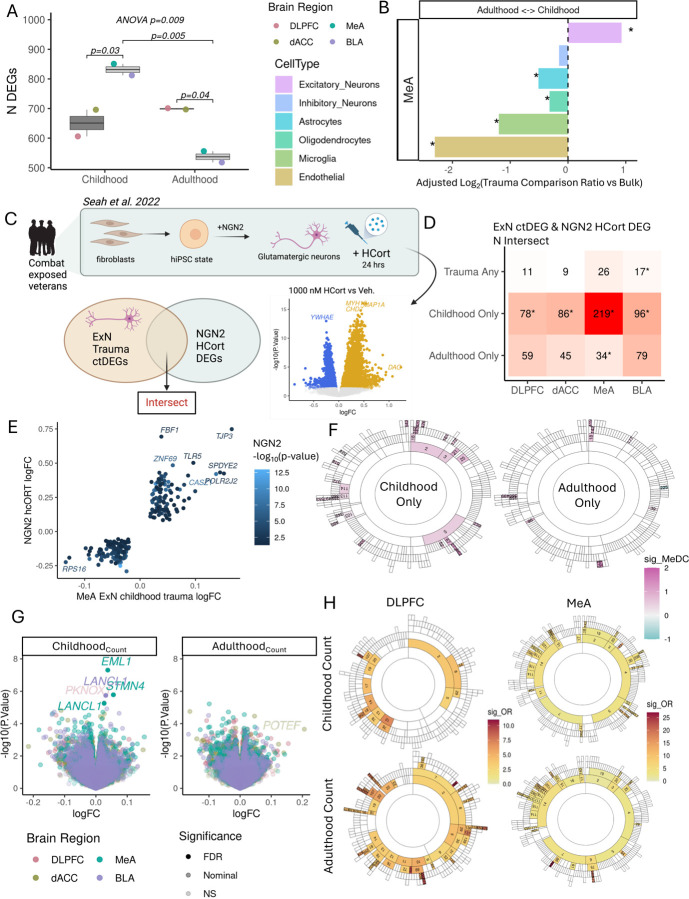
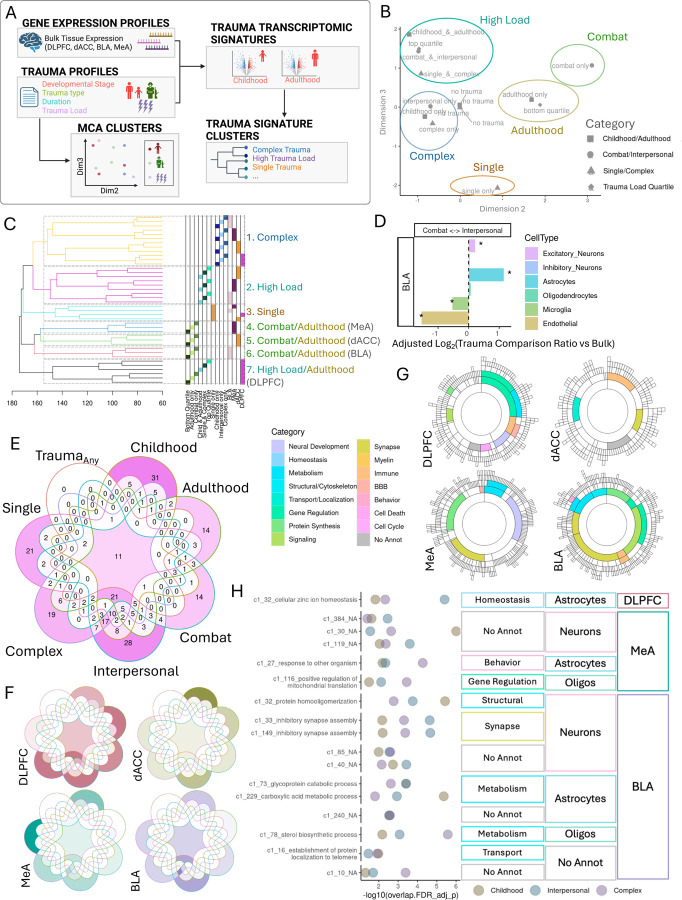
Psychological trauma has profound effects on brain function and precipitates psychiatric disorders in vulnerable individuals, however, the molecular mechanisms linking trauma with psychiatric risk remain incompletely understood. Using RNA-seq data postmortem brain tissue of a cohort of 304 donors (N=136 with trauma exposure), we investigated transcriptional signatures of trauma exposures in two cortical regions (dorsolateral prefrontal cortex, and dorsal anterior cingulate cortex) and two amygdala regions (medial amygdala and basolateral amygdala) associated with stress processing and regulation. We focused on dissecting heterogeneity of traumatic experiences in these transcriptional signatures by investigating exposure to several trauma types (childhood, adulthood, complex, single acute, combat, and interpersonal traumas) and interactions with sex. Overall, amygdala regions were more vulnerable to childhood traumas, whereas cortical regions were more vulnerable to adulthood trauma (regardless of childhood experience). Using cell-type-specific expression imputation, we identified a strong transcriptional response of medial amygdala excitatory neurons to childhood trauma, which coincided with dysregulation observed in a human induced pluripotent stem cell (hiPSC)-derived glutamatergic neurons exposed to hydrocortisone. We resolved multiscale coexpression networks for each brain region and identified modules enriched in trauma signatures and whose connectivity was altered with trauma. Trauma-associated coexpression modules provide insight into coordinated functional dysregulation with different traumas and point to potential gene targets for further dissection. Together, these data provide a characterization of the long-lasting human encoding of traumatic experiences in corticolimbic regions of human brain.
Conflict of interest statement
CONFLICT STATEMENT J.H.K. has consulting agreements (less than US$10,000 per year) with the following: Aptinyx, Inc. Biogen, Idec, MA, Bionomics, Limited (Australia), Boehringer Ingelheim International, Epiodyne, Inc., EpiVario, Inc., Janssen Research & Development, Jazz Pharmaceuticals, Inc., Otsuka America Pharmaceutical, Inc., Spring Care, Inc., Sunovion Pharmaceuticals, Inc.; is the co-founder for Freedom Biosciences, Inc.; serves on the scientific advisory boards of Biohaven Pharmaceuticals, BioXcel Therapeutics, Inc. (Clinical Advisory Board), Cerevel Therapeutics, LLC, Delix Therapeutics, Inc., Eisai, Inc., EpiVario, Inc., Jazz Pharmaceuticals, Inc., Neumora Therapeutics, Inc., Neurocrine Biosciences, Inc., Novartis Pharmaceuticals Corporation, PsychoGenics, Inc., Takeda Pharmaceuticals, Tempero Bio, Inc., Terran Biosciences, Inc..; has stock options with Biohaven Pharmaceuticals Medical Sciences, Cartego Therapeutics, Damona Pharmaceuticals, Delix Therapeutics, EpiVario, Inc., Neumora Therapeutics, Inc., Rest Therapeutics, Tempero Bio, Inc., Terran Biosciences, Inc., Tetricus, Inc.; and is editor of Biological Psychiatry with income greater than $10,000.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources