

This is a preprint.
A rapid, facile, and economical method for the isolation of ribosomes and translational machinery for structural and functional studies
- PMID: 39484553
- PMCID: PMC11526893
- DOI: 10.1101/2024.10.21.619433
A rapid, facile, and economical method for the isolation of ribosomes and translational machinery for structural and functional studies
Update in
-
A rapid, simple, and economical method for the isolation of ribosomes and translational machinery for structural and functional studies.Nat Commun. 2025 Aug 5;16(1):7185. doi: 10.1038/s41467-025-62314-8. Nat Commun. 2025. PMID: 40764614 Free PMC article.
Abstract
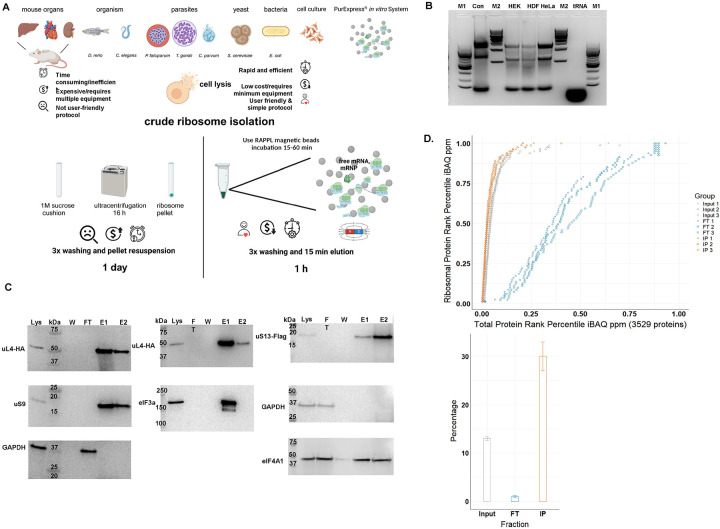
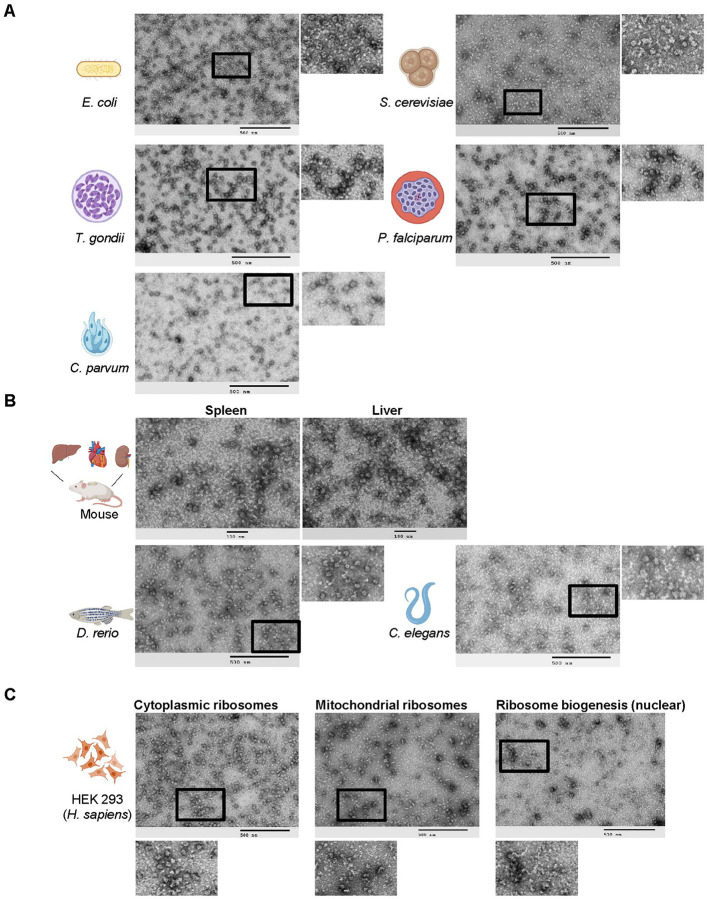
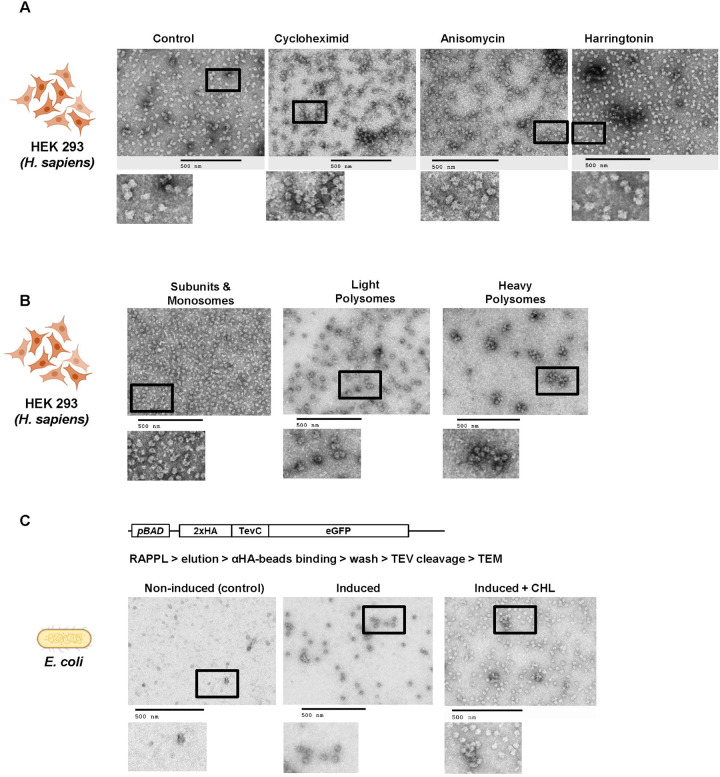
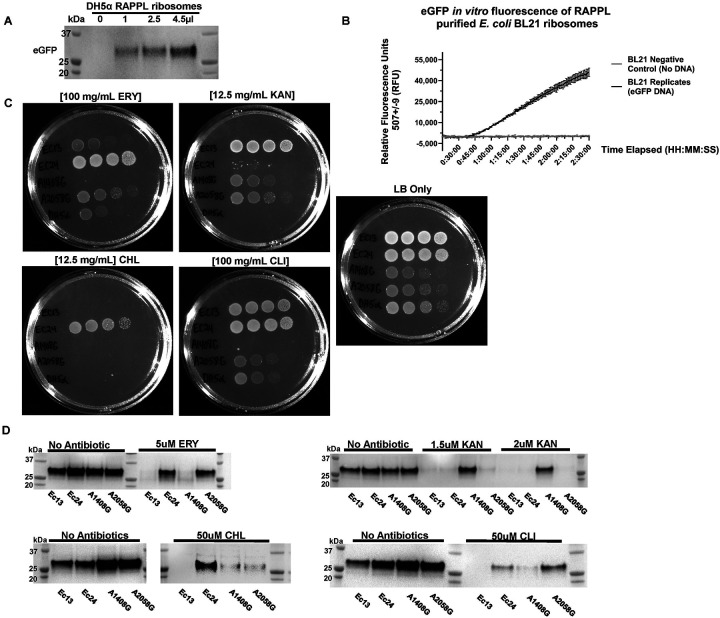
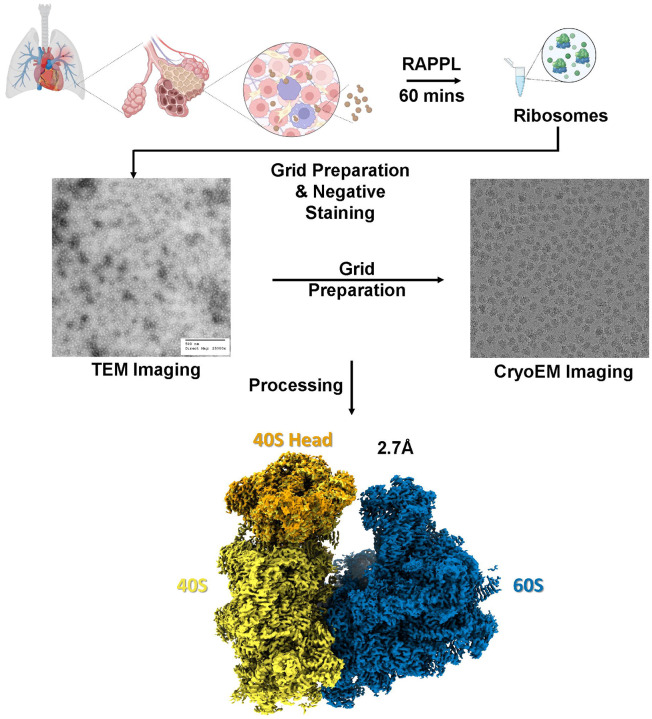
Ribosomes are macromolecular RNA-protein complexes that constitute the central machinery responsible for protein synthesis and quality control in the cell. Ribosomes also serve as a hub for multiple non-ribosomal proteins and RNAs that control protein synthesis. However, the purification of ribosomes and associated factors for functional and structural studies requires a large amount of starting biological material and a tedious workflow. Current methods are challenging as they combine ultracentrifugation, the use of sucrose cushions or gradients, expensive equipment, and multiple hours to days of work. Here, we present a rapid, facile, and cost-effective method to isolate ribosomes from in vivo or in vitro samples for functional and structural studies using single-step enrichment on magnetic beads - RAPPL (RNA Affinity Purification using Poly-Lysine). Using mass spectrometry and western blot analyses, we show that poly-lysine coated beads incubated with E. coli and HEK-293 cell lysates enrich specifically for ribosomes and ribosome-associated factors. We demonstrate the ability of RAPPL to isolate ribosomes and translation-associated factors from limited material quantities, as well as a wide variety of biological samples: cell lysates, cells, organs, and whole organisms. Using RAPPL, we characterized and visualized the different effects of various drugs and translation inhibitors on protein synthesis. Our method is compatible with traditional ribosome isolation. It can be used to purify specific complexes from fractions of sucrose gradients or in tandem affinity purifications for ribosome-associated factors. Ribosomes isolated using RAPPL are functionally active and can be used for rapid screening and in vitro characterization of ribosome antibiotic resistance. Lastly, we demonstrate the structural applications of RAPPL by purifying and solving the 2.7Å cryo-EM structure of ribosomes from the Cryptococcus neoformans, an encapsulated yeast causing cryptococcosis. Ribosomes and translational machinery purified with this method are suitable for subsequent functional or structural analyses and provide a solid foundation for researchers to carry out further applications - academic, clinical, or industrial - on ribosomes.
Figures

References
-
- Keller E. B., Zamecnik P. C., Loftfield R. B., The role of microsomes in the incorporation of amino acids into proteins. J Histochem Cytochem 2, 378–386 (1954). - PubMed
-
- Ban N., Nissen P., Hansen J., Moore P. B., Steitz T. A., The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science 289, 905–920 (2000). - PubMed
-
- Moore P. B., Steitz T. A., The structural basis of large ribosomal subunit function. Annu Rev Biochem 72, 813–850 (2003). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources