

Prostaglandin E2-EP2/EP4 signaling induces immunosuppression in human cancer by impairing bioenergetics and ribosome biogenesis in immune cells
- PMID: 39487111
- PMCID: PMC11530437
- DOI: 10.1038/s41467-024-53706-3
Prostaglandin E2-EP2/EP4 signaling induces immunosuppression in human cancer by impairing bioenergetics and ribosome biogenesis in immune cells
Abstract
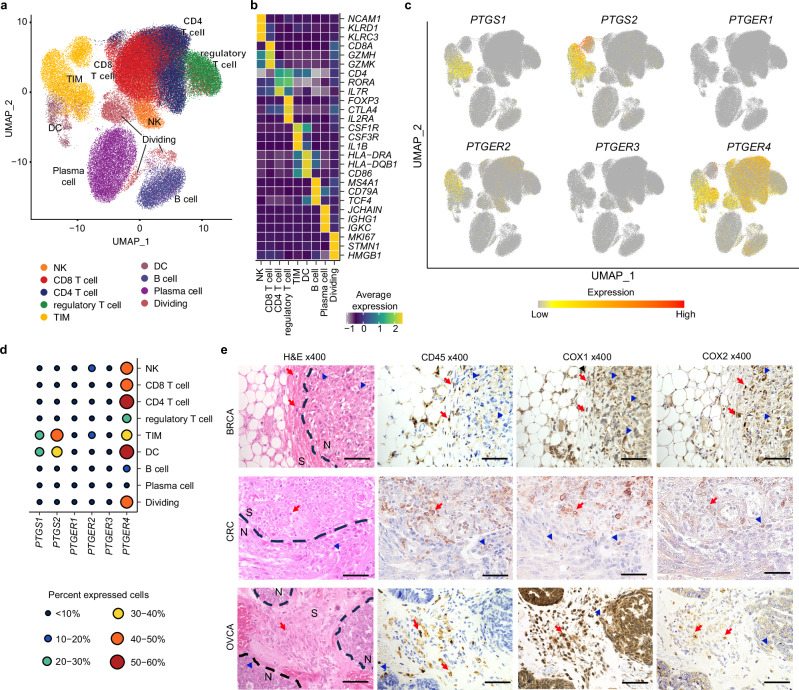
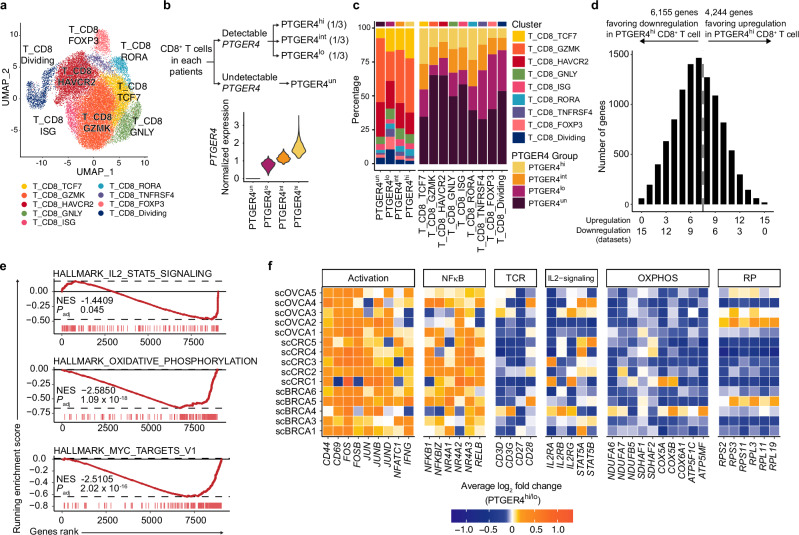
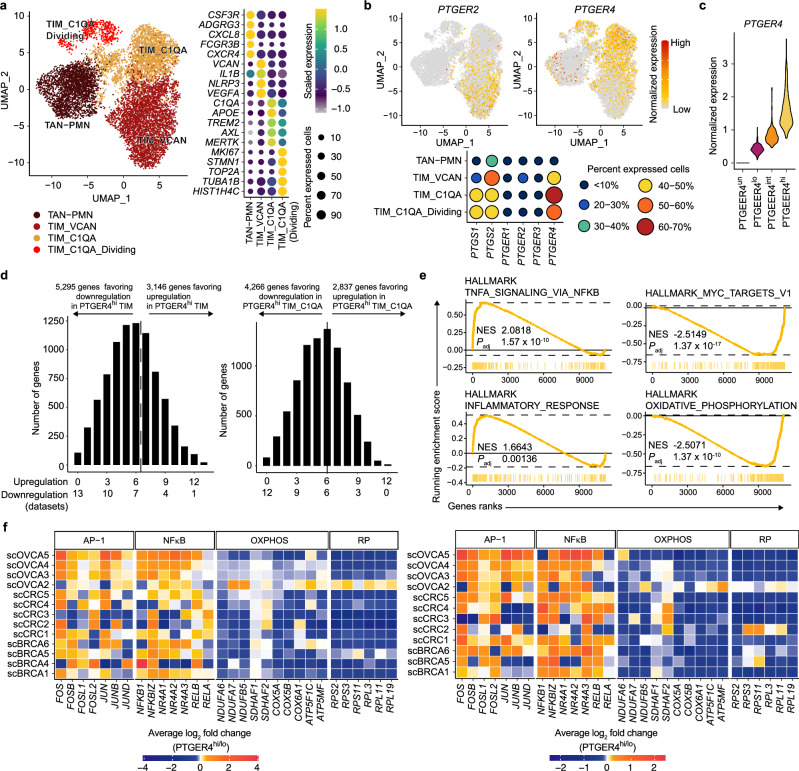
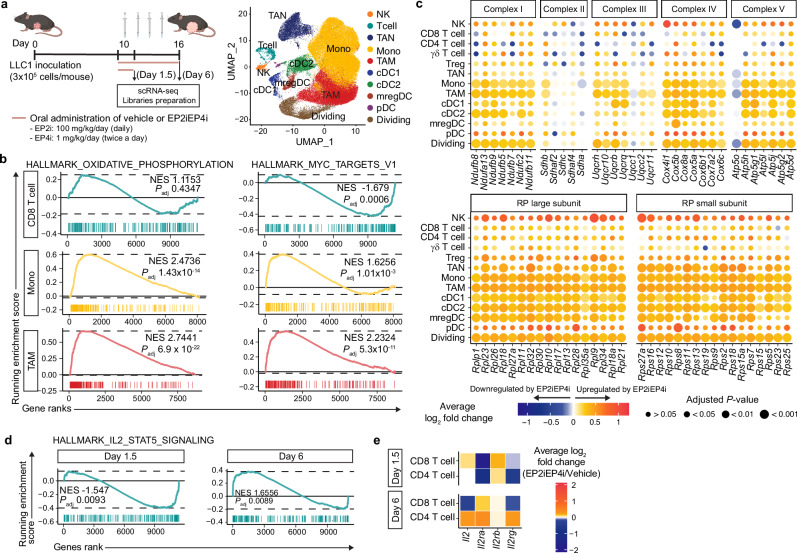
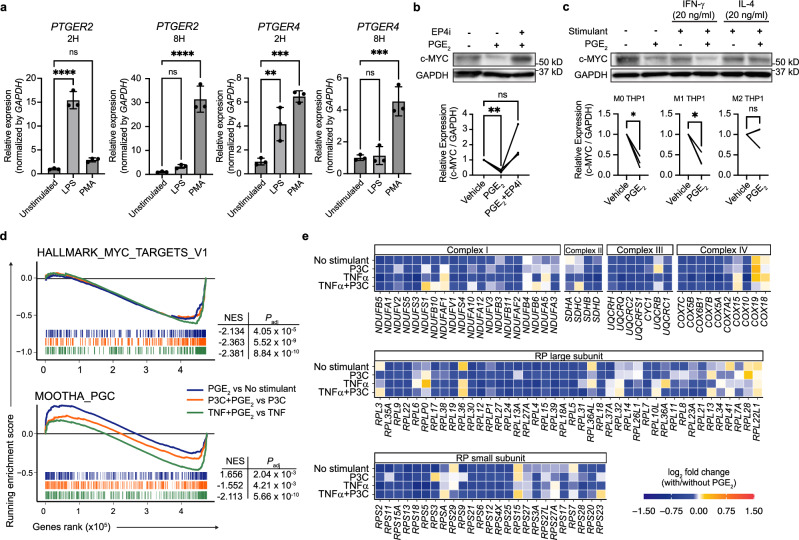
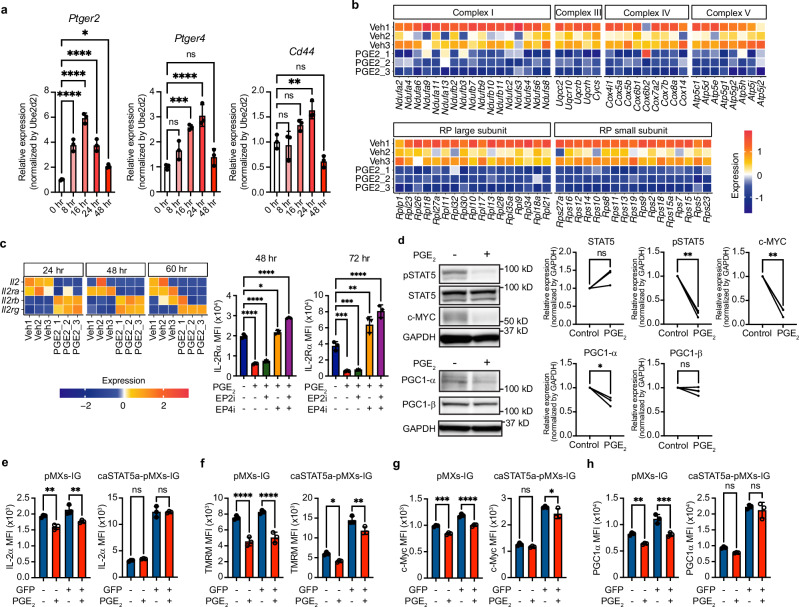
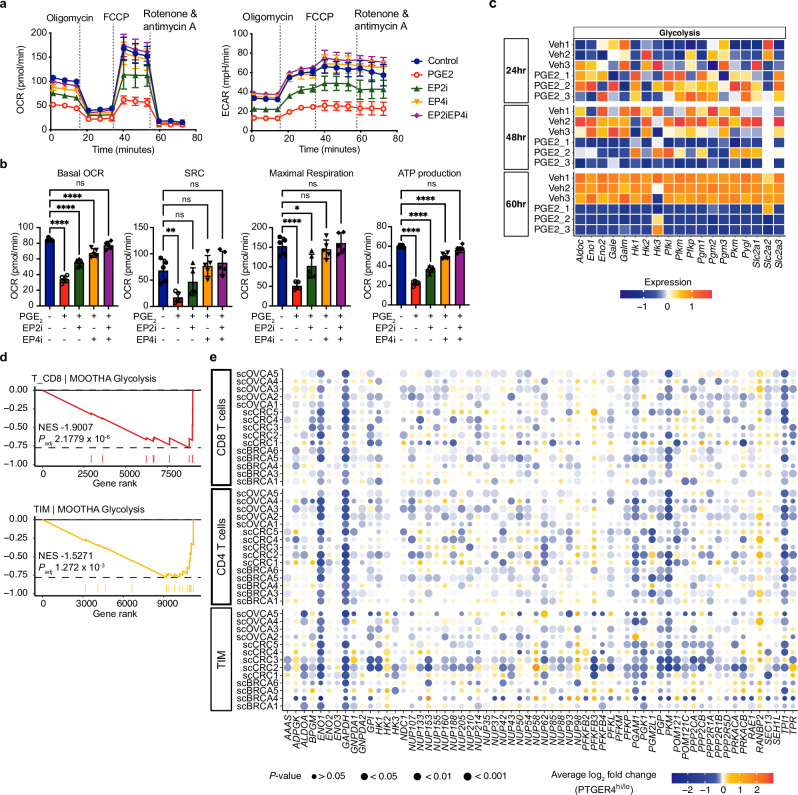
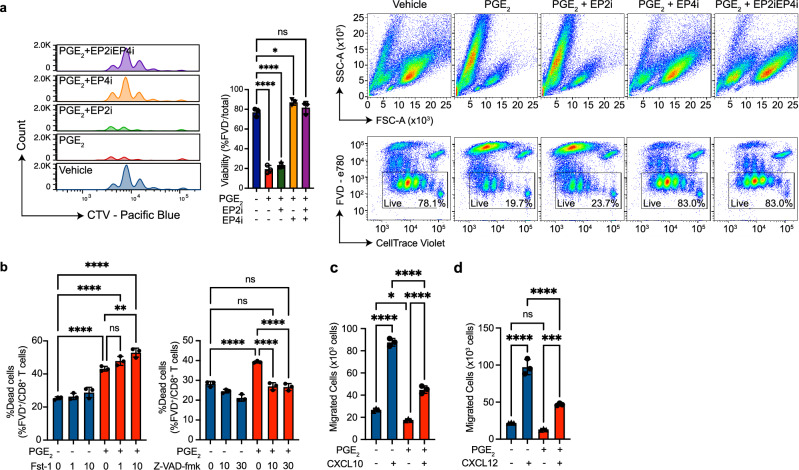
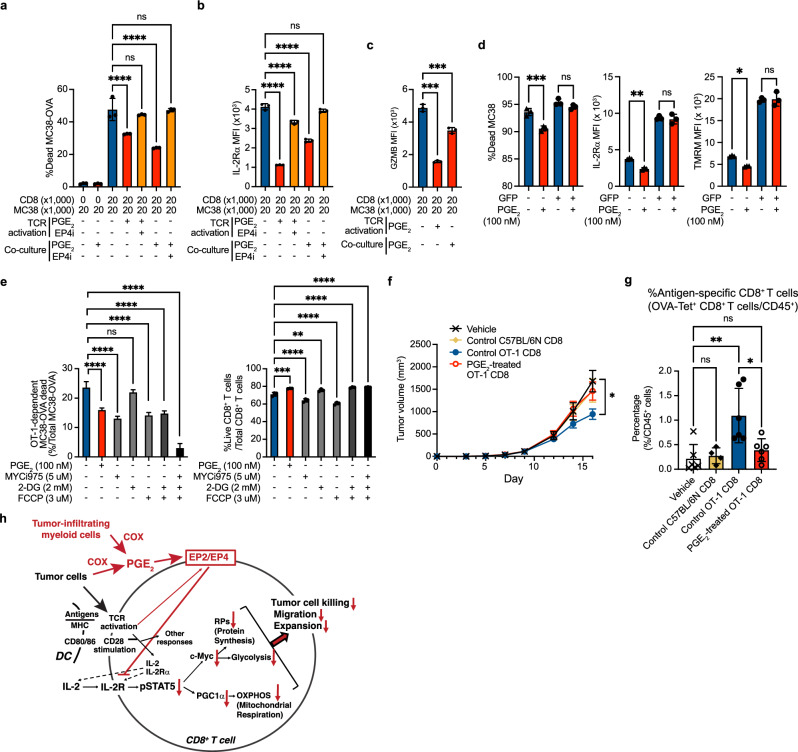
While prostaglandin E2 (PGE2) is produced in human tumor microenvironment (TME), its role therein remains poorly understood. Here, we examine this issue by comparative single-cell RNA sequencing of immune cells infiltrating human cancers and syngeneic tumors in female mice. PGE receptors EP4 and EP2 are expressed in lymphocytes and myeloid cells, and their expression is associated with the downregulation of oxidative phosphorylation (OXPHOS) and MYC targets, glycolysis and ribosomal proteins (RPs). Mechanistically, CD8+ T cells express EP4 and EP2 upon TCR activation, and PGE2 blocks IL-2-STAT5 signaling by downregulating Il2ra, which downregulates c-Myc and PGC-1 to decrease OXPHOS, glycolysis, and RPs, impairing migration, expansion, survival, and antitumor activity. Similarly, EP4 and EP2 are induced upon macrophage activation, and PGE2 downregulates c-Myc and OXPHOS in M1-like macrophages. These results suggest that PGE2-EP4/EP2 signaling impairs both adaptive and innate immunity in TME by hampering bioenergetics and ribosome biogenesis of tumor-infiltrating immune cells.
© 2024. The Author(s).
Conflict of interest statement
S.N. is a scientific advisor to Astellas Pharma Inc. and Toray Co Ltd. This work was supported in part by the Collaborative Research Grant from Ono Pharmaceutical Co Ltd. The remaining authors declare no competing interests.
Figures
References
-
- Rothwell, P. M. et al. Effect of daily aspirin on long-term risk of death due to cancer: analysis of individual patient data from randomised trials. Lancet77, 31–41 (2011). - PubMed
-
- Riciotti, E., Grosser, T. & FitzGerald, G. A. Lipid-derived autacoids: eicosanoids and platelet-activating factor in Goodman & Gilman’s The Pharmacological Basis of Therapeutics 14th edn (eds Brunton, L. L. & Knollmann, B. C.) 813–828 (McGraw-Hill, 2023).
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
