

Transfer of mitochondrial DNA into the nuclear genome during induced DNA breaks
- PMID: 39487167
- PMCID: PMC11530683
- DOI: 10.1038/s41467-024-53806-0
Transfer of mitochondrial DNA into the nuclear genome during induced DNA breaks
Abstract
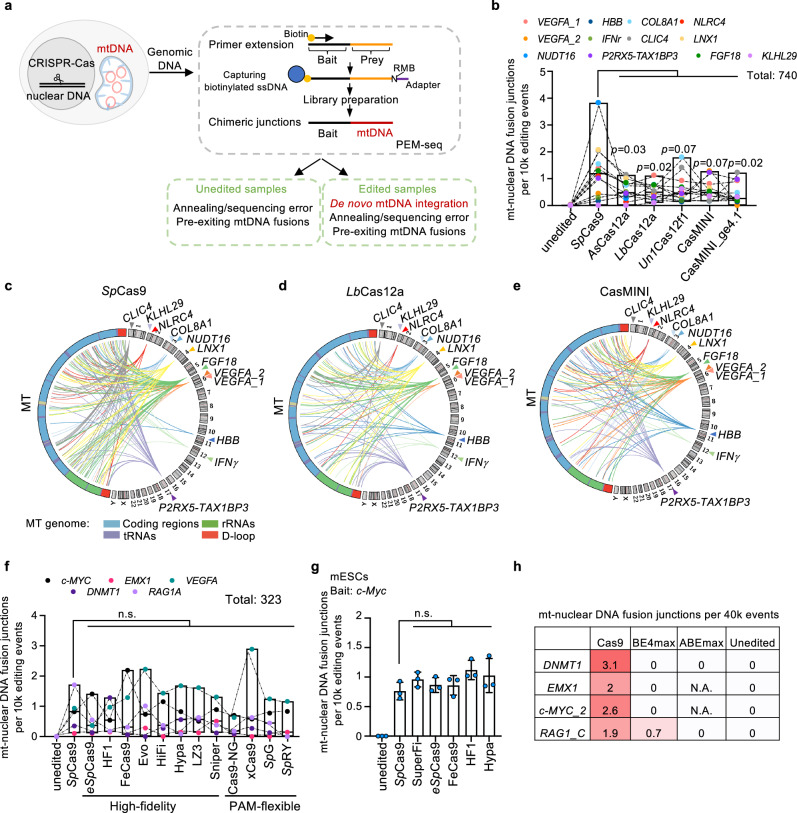
Mitochondria serve as the cellular powerhouse, and their distinct DNA makes them a prospective target for gene editing to treat genetic disorders. However, the impact of genome editing on mitochondrial DNA (mtDNA) stability remains a mystery. Our study reveals previously unknown risks of genome editing that both nuclear and mitochondrial editing cause discernible transfer of mitochondrial DNA segments into the nuclear genome in various cell types including human cell lines, primary T cells, and mouse embryos. Furthermore, drug-induced mitochondrial stresses and mtDNA breaks exacerbate this transfer of mtDNA into the nuclear genome. Notably, we observe that mitochondrial editors, including mitoTALEN and recently developed base editor DdCBE, can also enhance crosstalk between mtDNA and the nuclear genome. Moreover, we provide a practical solution by co-expressing TREX1 or TREX2 exonucleases during DdCBE editing. These findings imply genome instability of mitochondria during induced DNA breaks and explain the origins of mitochondrial-nuclear DNA segments.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- SRA/SRP119022
- SRA/PRJNA786071
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
