

Lateral lamina V projection neuron axon collaterals connect sensory processing across the dorsal horn of the mouse spinal cord
- PMID: 39487174
- PMCID: PMC11530558
- DOI: 10.1038/s41598-024-73620-4
Lateral lamina V projection neuron axon collaterals connect sensory processing across the dorsal horn of the mouse spinal cord
Abstract
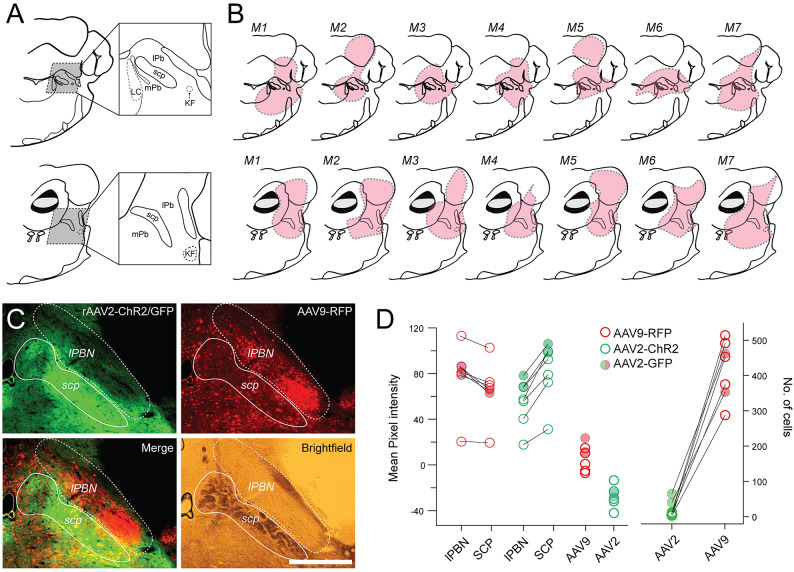
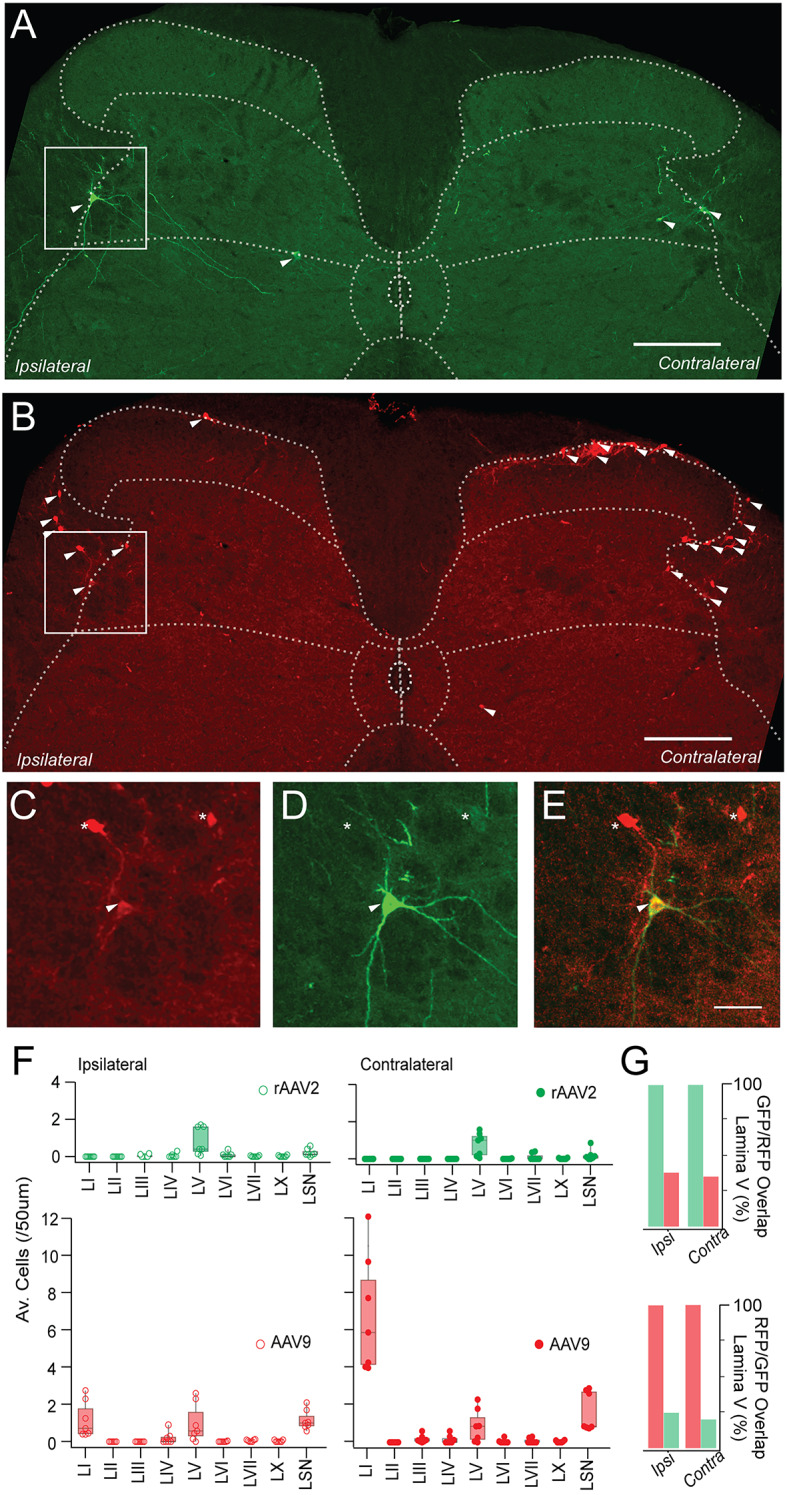
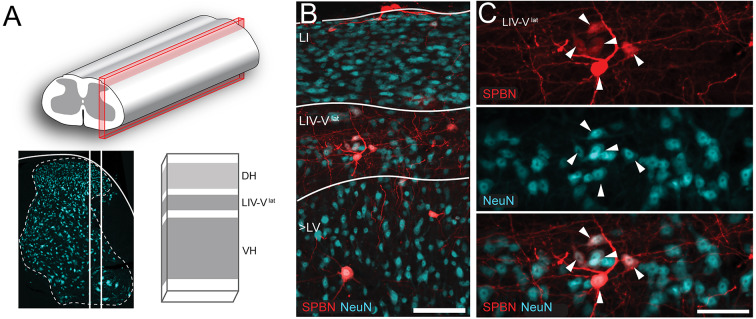
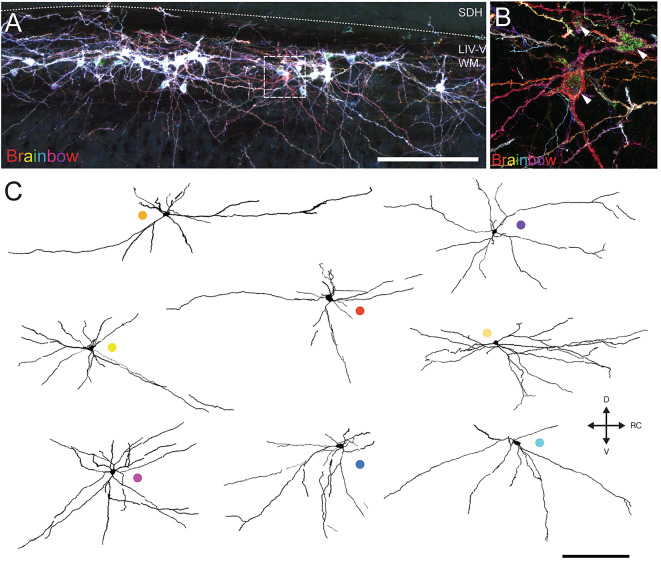
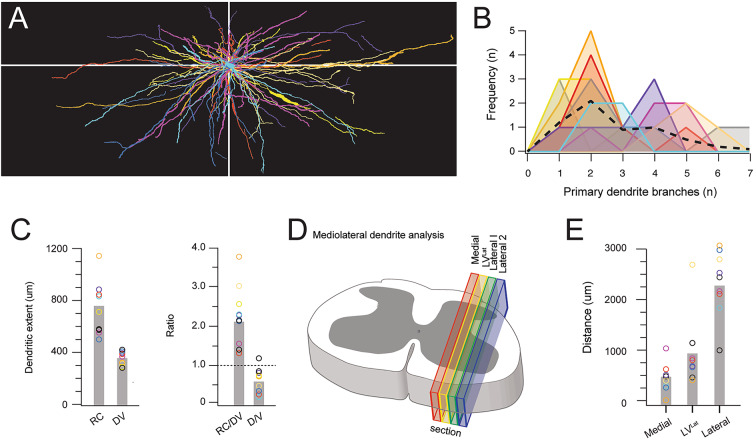
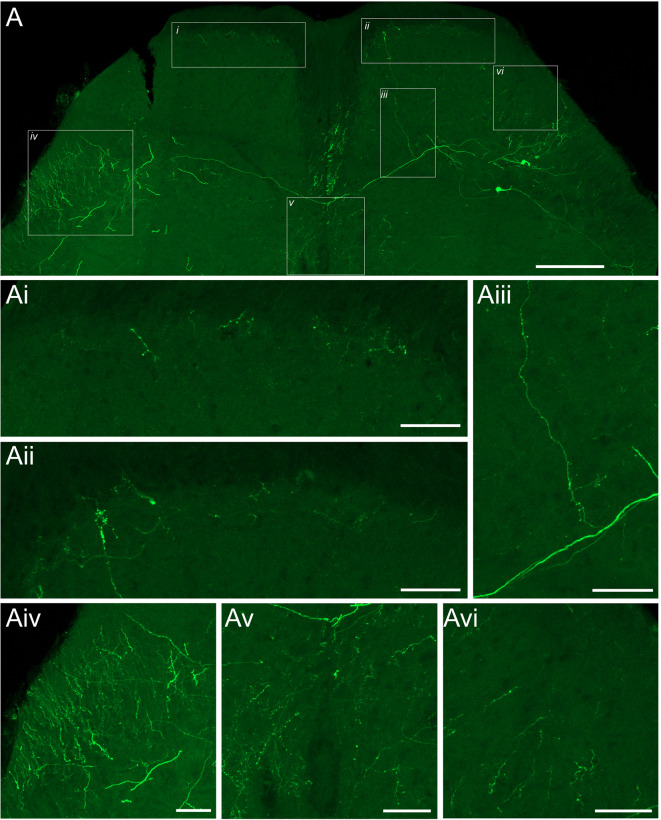
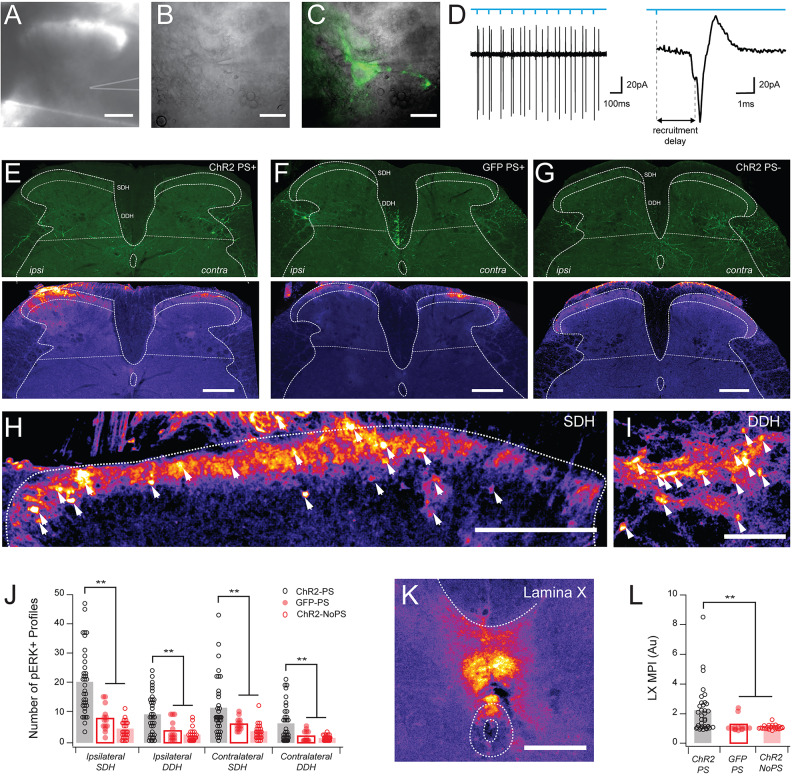
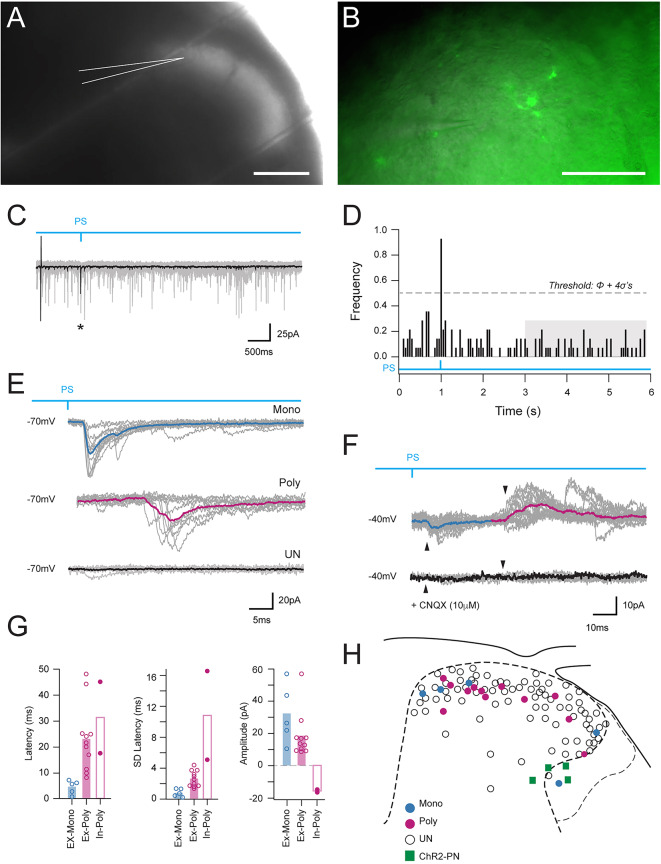
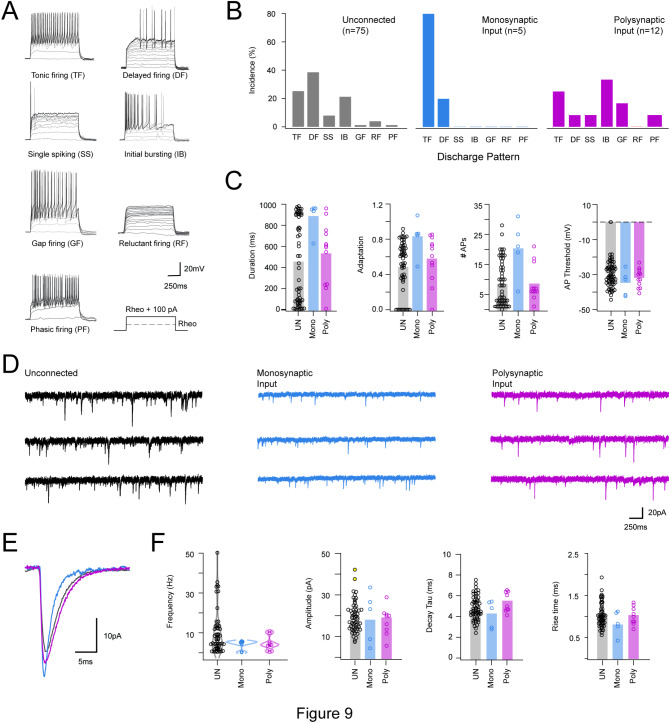
Spinal projection neurons (PNs) are defined by long axons that travel from their origin in the spinal cord to the brain where they relay sensory information from the body. The existence and function of a substantial axon collateral network, also arising from PNs and remaining within the spinal cord, is less well appreciated. Here we use a retrograde viral transduction strategy to characterise a novel subpopulation of deep dorsal horn spinoparabrachial neurons. Brainbow assisted analysis confirmed that virally labelled PN cell bodies formed a discrete cell column in the lateral part of Lamina V (LVlat) and the adjoining white matter. These PNs exhibited large dendritic territories biased to regions lateral and ventral to the cell body column and extending considerable rostrocaudal distances. Optogenetic activation of LVLat PNs confirmed this population mediates widespread signalling within spinal cord circuits, including activation in the superficial dorsal horn. This signalling was also demonstrated with patch clamp recordings during LVLat PN photostimulation, with a range of direct and indirect connections identified and evidence of a postsynaptic population of inhibitory interneurons. Together, these findings confirm a substantial role for PNs in local spinal sensory processing, as well as relay of sensory signals to the brain.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Carlton, S. M. Nociceptive primary afferents: they have a mind of their own. J. Physiol.592, 3403–3411. 10.1113/jphysiol.2013.269654 (2014). https://doi.org:. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
