

CLSTN3B promotes lipid droplet maturation and lipid storage in mouse adipocytes
- PMID: 39488519
- PMCID: PMC11531554
- DOI: 10.1038/s41467-024-53750-z
CLSTN3B promotes lipid droplet maturation and lipid storage in mouse adipocytes
Abstract
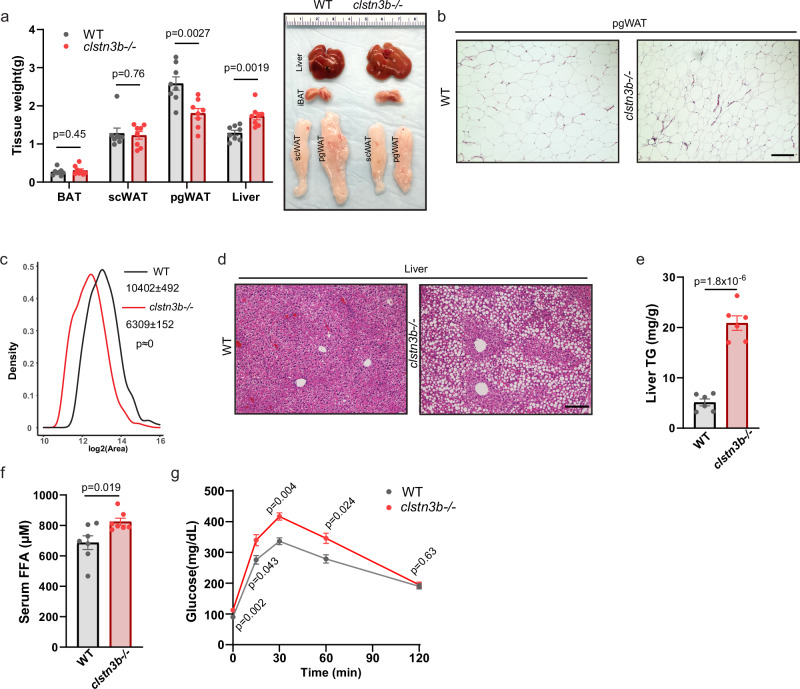
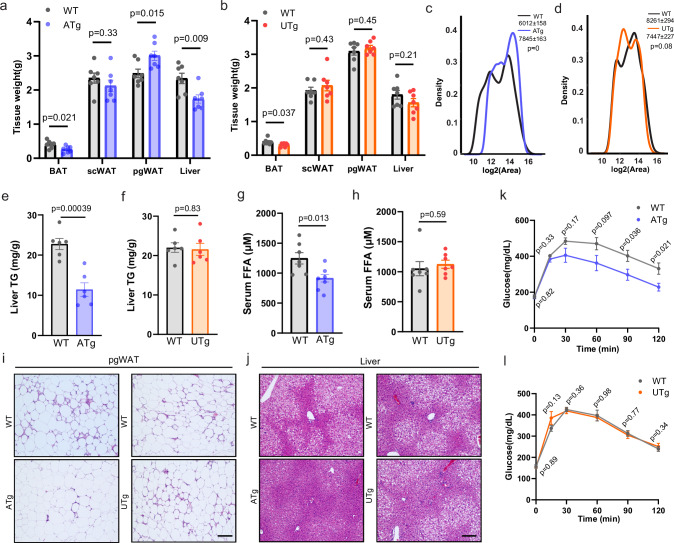
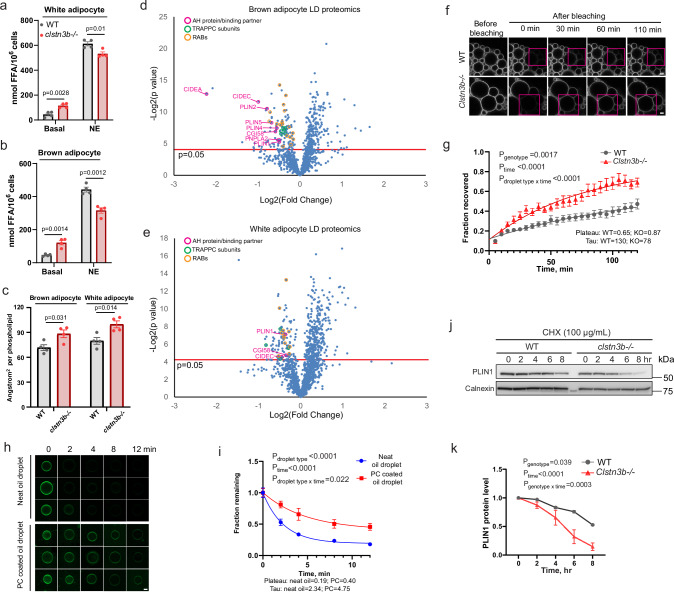
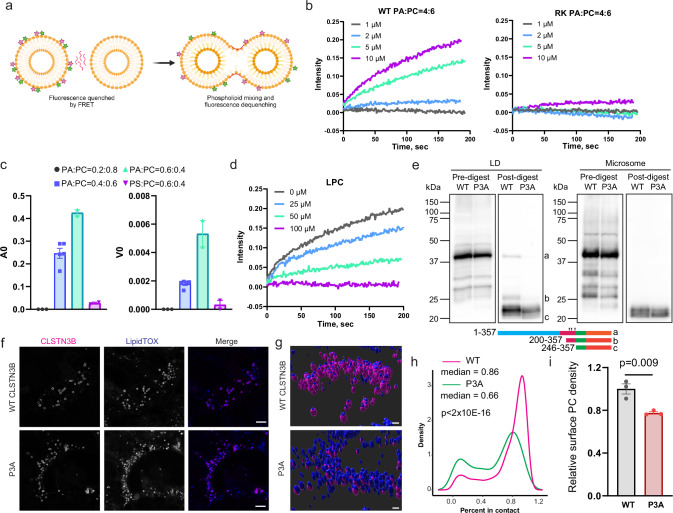
Interorganelle contacts facilitate material exchanges and sustain the structural and functional integrity of organelles. Lipid droplets (LDs) of adipocytes are responsible for energy storage and mobilization responding to body needs. LD biogenesis defects compromise the lipid-storing capacity of adipocytes, resulting in ectopic lipid deposition and metabolic disorders, yet how the uniquely large LDs in adipocytes attain structural and functional maturation is incompletely understood. Here we show that the mammalian adipocyte-specific protein CLSTN3B is crucial for adipocyte LD maturation. CLSTN3B employs an arginine-rich segment to promote extensive contact and hemifusion-like structure formation between the endoplasmic reticulum (ER) and LD, allowing ER-to-LD phospholipid diffusion during LD expansion. CLSTN3B ablation results in reduced LD surface phospholipid density, increased turnover of LD-surface proteins, and impaired LD functions. Our results establish the central role of CLSTN3B in the adipocyte-specific LD maturation pathway that enhances lipid storage and maintenance of metabolic health under caloric overload in mice of both sexes.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
CLSTN3B enhances adipocyte lipid droplet structure and function via endoplasmic reticulum contact.bioRxiv [Preprint]. 2024 Jan 21:2024.01.20.576491. doi: 10.1101/2024.01.20.576491. bioRxiv. 2024. PMID: 38293096 Free PMC article. Preprint.
-
CLSTN3β enforces adipocyte multilocularity to facilitate lipid utilization.Nature. 2023 Jan;613(7942):160-168. doi: 10.1038/s41586-022-05507-1. Epub 2022 Dec 7. Nature. 2023. PMID: 36477540 Free PMC article.
-
ORP5 and ORP8 orchestrate lipid droplet biogenesis and maintenance at ER-mitochondria contact sites.J Cell Biol. 2022 Sep 5;221(9):e202112107. doi: 10.1083/jcb.202112107. Epub 2022 Aug 12. J Cell Biol. 2022. PMID: 35969857 Free PMC article.
-
Lipid Droplets Big and Small: Basic Mechanisms That Make Them All.Annu Rev Cell Dev Biol. 2024 Oct;40(1):143-168. doi: 10.1146/annurev-cellbio-012624-031419. Annu Rev Cell Dev Biol. 2024. PMID: 39356808 Review.
-
Born this way - Biogenesis of lipid droplets from specialized ER subdomains.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Jan;1865(1):158448. doi: 10.1016/j.bbalip.2019.04.008. Epub 2019 Apr 24. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31028912 Review.
Cited by
-
PPARα regulates ER-lipid droplet protein Calsyntenin-3β to promote ketogenesis in hepatocytes.Proc Natl Acad Sci U S A. 2025 Apr 29;122(17):e2426338122. doi: 10.1073/pnas.2426338122. Epub 2025 Apr 21. Proc Natl Acad Sci U S A. 2025. PMID: 40258152
-
Hepatocyte-specific CLSTN3B ablation impairs lipid droplet maturation and alleviates diet-induced steatohepatitis in mice.bioRxiv [Preprint]. 2025 May 25:2025.05.20.655199. doi: 10.1101/2025.05.20.655199. bioRxiv. 2025. PMID: 40475430 Free PMC article. Preprint.
-
Porcine SCD1 Regulates Lipid Droplet Number via CLSTN3B in PK15 Cells.Animals (Basel). 2025 Jun 4;15(11):1663. doi: 10.3390/ani15111663. Animals (Basel). 2025. PMID: 40509129 Free PMC article.
References
-
- Shulman, G. I. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med.371, 1131–1141 (2014). - PubMed
-
- Akazawa, S., Sun, F., Ito, M., Kawasaki, E. & Eguchi, K. Efficacy of troglitazone on body fat distribution in type 2 diabetes. Diab. Care23, 1067–1071 (2000). - PubMed
-
- Yki-Järvinen, H. Thiazolidinediones. N. Engl. J. Med.351, 1106–1118 (2004). - PubMed
-
- Lipscombe, L. L. et al. Thiazolidinediones and cardiovascular outcomes in older patients with diabetes. JAMA298, 2634–2643 (2007). - PubMed
MeSH terms
Substances
Grants and funding
- R01 DK135556/DK/NIDDK NIH HHS/United States
- 23CDA1050474/American Heart Association (American Heart Association, Inc.)
- RR200084/Cancer Prevention and Research Institute of Texas (Cancer Prevention Research Institute of Texas)
- R01DK135556/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
LinkOut - more resources
Full Text Sources
Research Materials
