

The role of mesenchymal cells in cholangiocarcinoma
- PMID: 39492622
- PMCID: PMC11655028
- DOI: 10.1242/dmm.050716
The role of mesenchymal cells in cholangiocarcinoma
Abstract
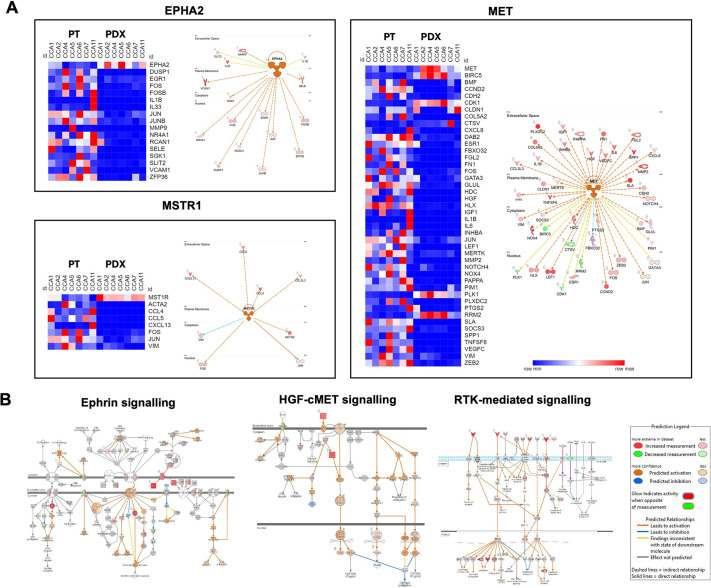
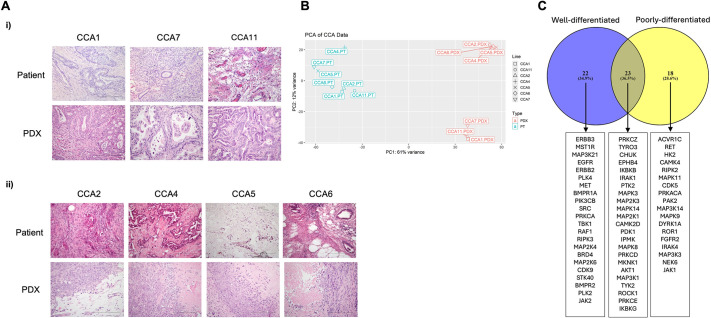
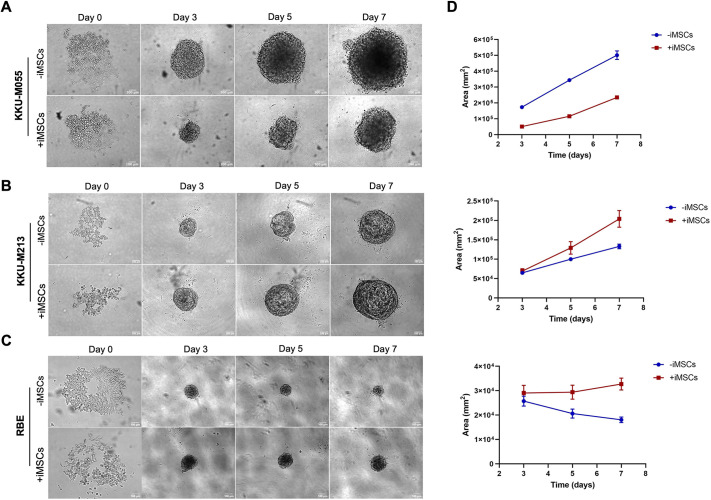
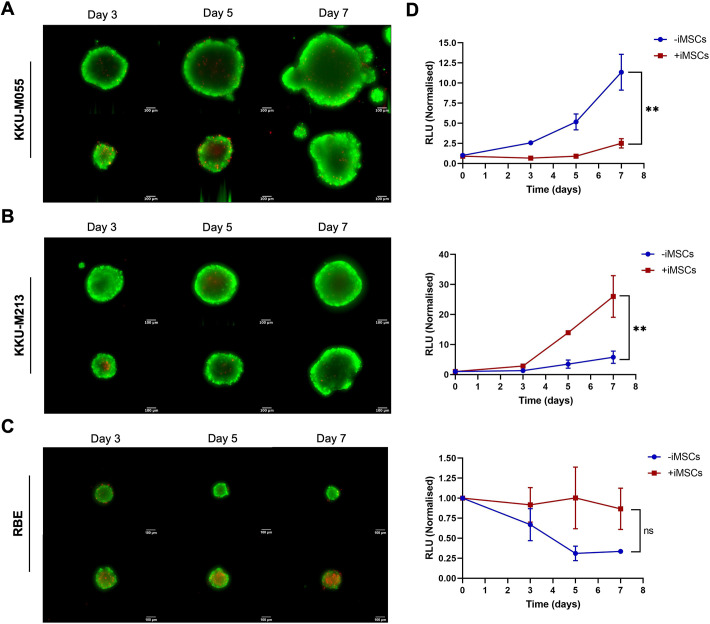
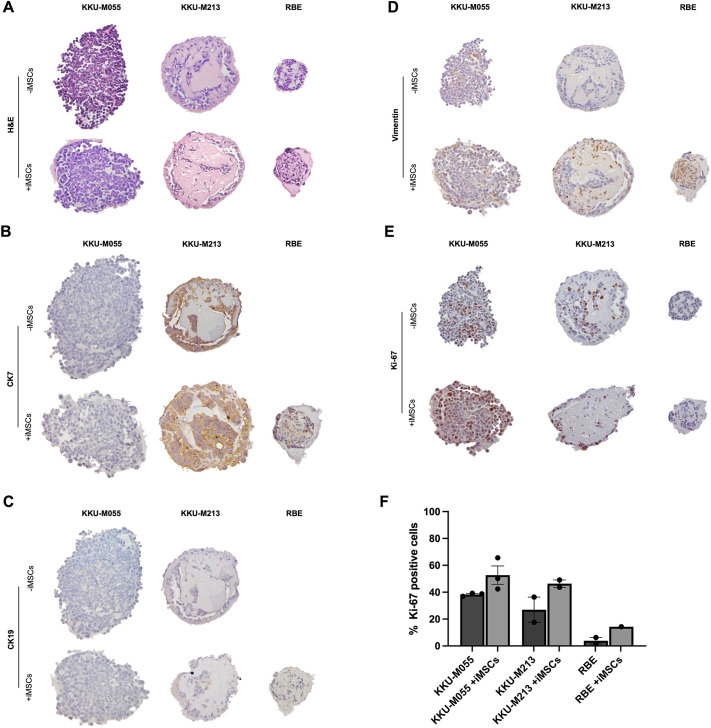
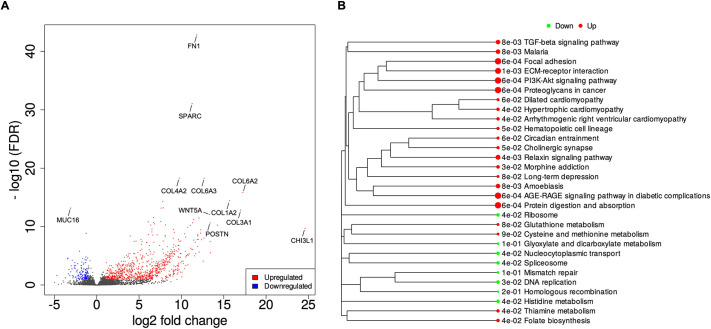
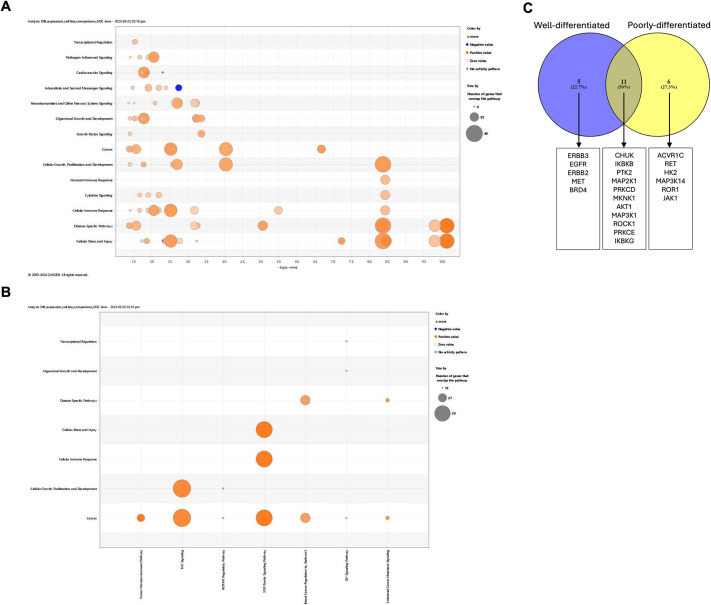
The tumour microenvironment (TME) significantly influences tumour formation and progression through dynamic interactions. Cholangiocarcinoma (CCA), a highly desmoplastic tumour, lacks early diagnostic biomarkers and has limited effective treatments owing to incomplete understanding of its molecular pathogenesis. Investigating the role of the TME in CCA progression could lead to better therapies. RNA sequencing was performed on seven CCA patient-derived xenografts (PDXs) and their corresponding patient samples. Differential expression analysis was conducted, and Qiagen Ingenuity Pathway Analysis was used to predict dysregulated pathways and upstream regulators. PDX- and cell line-derived spheroids, with and without immortalised mesenchymal stem cells, were grown and analysed for morphology, growth and viability. Histological analysis confirmed biliary phenotypes. RNA sequencing indicated upregulation of extracellular matrix-receptor interaction and PI3K-AKT pathways in the presence of mesenchymal cells, with several genes linked to poor survival. Mesenchymal cells restored the activity of inhibited cancer-associated kinases. Thus, adding mesenchymal cells to CCA spheroid models restored key paracrine signalling pathways lost in PDXs, enhancing tumour growth and viability. These findings highlight the importance of including stromal components in cancer models to improve pre-clinical studies.
Keywords: Cholangiocarcinoma; Mesenchymal stem cells; PDX models; Signalling pathways; Tumour microenvironment.
© 2024. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures

References
-
- Affo, S., Nair, A., Brundu, F., Ravichandra, A., Bhattacharjee, S., Matsuda, M., Chin, L., Filliol, A., Wen, W., Song, X.et al. (2021). Promotion of cholangiocarcinoma growth by diverse cancer-associated fibroblast subpopulations. Cancer Cell 39, 866-882.e11. 10.1016/j.ccell.2021.03.012 - DOI - PMC - PubMed
-
- Allen, I. C., Eden, K., Heid, B. and Holl, E. K. (2017). Map3K14 signaling attenuates the development of colorectal cancer through activation of the non-canonical NF-κB signaling cascade. J. Immunol. 198, 197.6-197.6. 10.4049/jimmunol.198.Supp.197.6 - DOI
-
- Banales, J. M., Marin, J. J. G., Lamarca, A., Rodrigues, M., Khan, S. A., Roberts, L. R., Cardinale, V., Carpino, G., Andersen, J. B., Braconi, C.et al. (2020). Cholangiocarcinoma 2020: the next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 17, 557-588. 10.1038/s41575-020-0310-z - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
