

Identification and characterization of the lipoprotein N-acyltransferase in Bacteroides
- PMID: 39495918
- PMCID: PMC11573676
- DOI: 10.1073/pnas.2410909121
Identification and characterization of the lipoprotein N-acyltransferase in Bacteroides
Abstract
Members of the Bacteroidota compose a large portion of the human gut microbiota, contributing to overall gut health via the degradation of various polysaccharides. This process is facilitated by lipoproteins, globular proteins anchored to the cell surface by a lipidated N-terminal cysteine. Despite their importance, lipoprotein synthesis by these bacteria is understudied. In Escherichia coli, the α-amino-linked lipid of lipoproteins is added by the
Keywords: Bacteroides; acyltransferase; cell surface; lipoproteins.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures

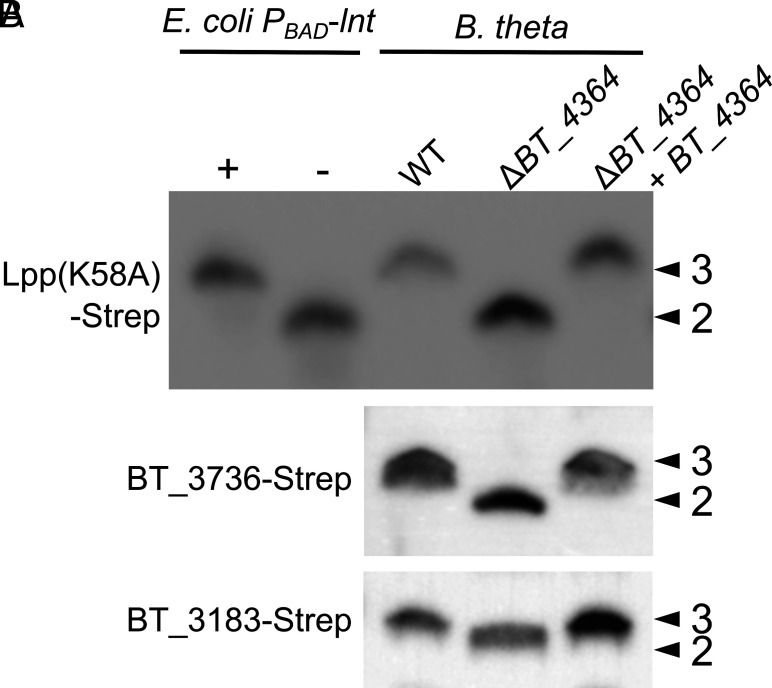

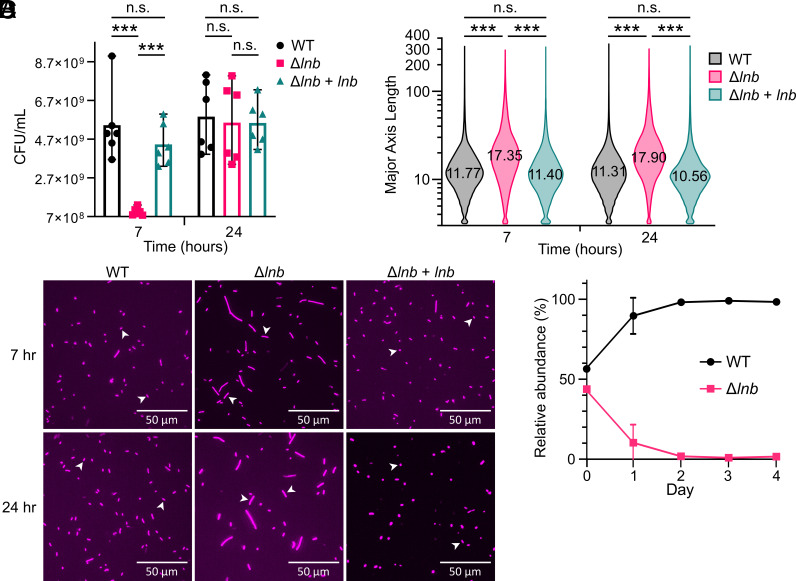
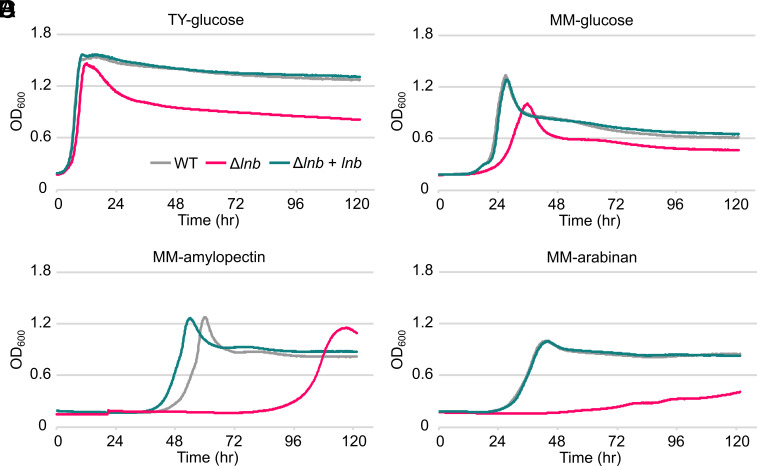


Update of
-
Identification and Characterization of the Lipoprotein N-acyltransferase in Bacteroides.bioRxiv [Preprint]. 2024 Jun 1:2024.05.31.596883. doi: 10.1101/2024.05.31.596883. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Nov 12;121(46):e2410909121. doi: 10.1073/pnas.2410909121. PMID: 38853980 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- R01DK125445/HHS | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK)
- R01 DK125445/DK/NIDDK NIH HHS/United States
- R21 AI180287/AI/NIAID NIH HHS/United States
- R21AI151873/HHS | NIH | NIAID | Division of Microbiology and Infectious Diseases (DMID)
- DP210100362/Department of Education and Training | Australian Research Council (ARC)
- R21 AI151873/AI/NIAID NIH HHS/United States
- R21 AI168719/AI/NIAID NIH HHS/United States
- R21AI168719/HHS | NIH | NIAID | Division of Microbiology and Infectious Diseases (DMID)
- N/A/Michigan Life Sciences Fellows Program
- FT200100270/Department of Education and Training | Australian Research Council (ARC)
- S10 OD034245/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
