

Neuroblastoma plasticity during metastatic progression stems from the dynamics of an early sympathetic transcriptomic trajectory
- PMID: 39500881
- PMCID: PMC11538482
- DOI: 10.1038/s41467-024-53776-3
Neuroblastoma plasticity during metastatic progression stems from the dynamics of an early sympathetic transcriptomic trajectory
Abstract
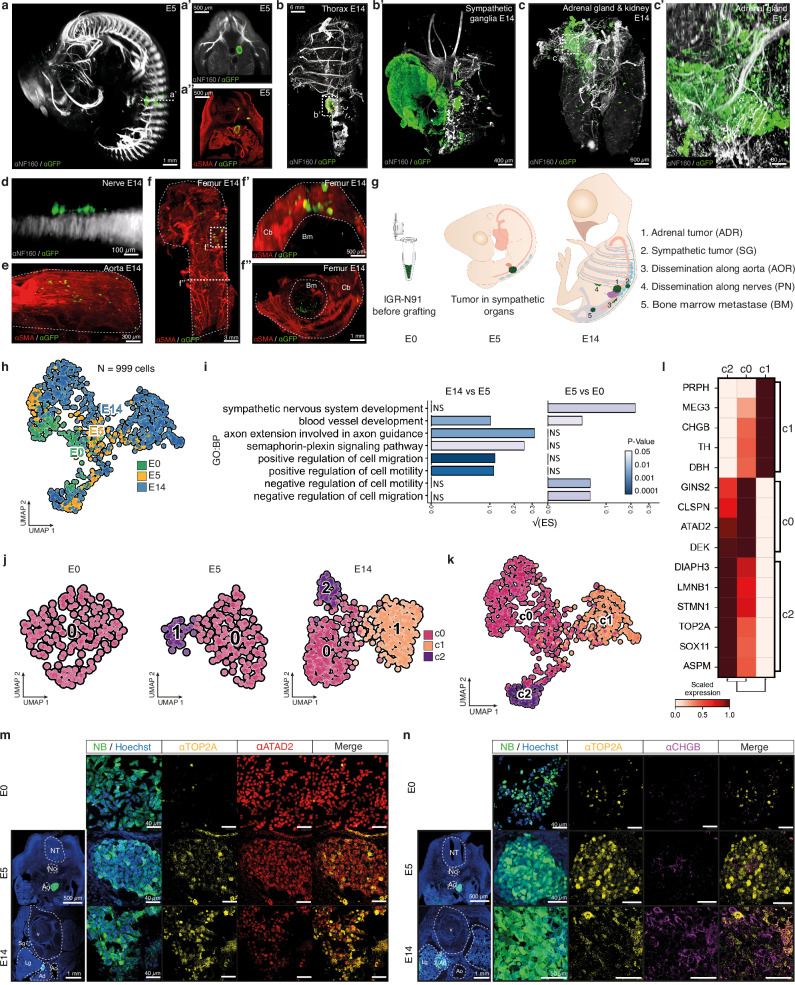
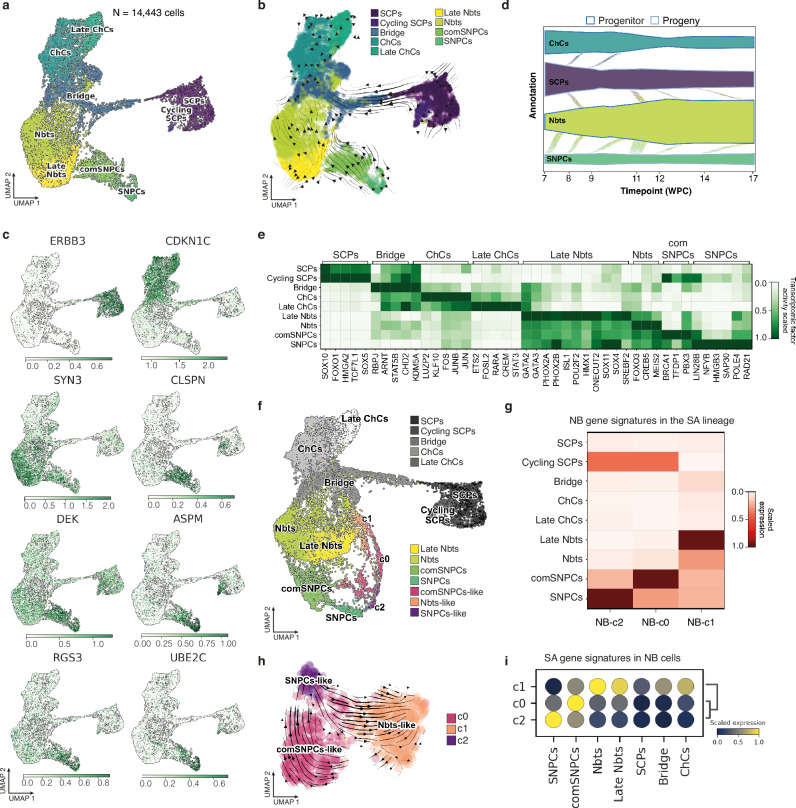
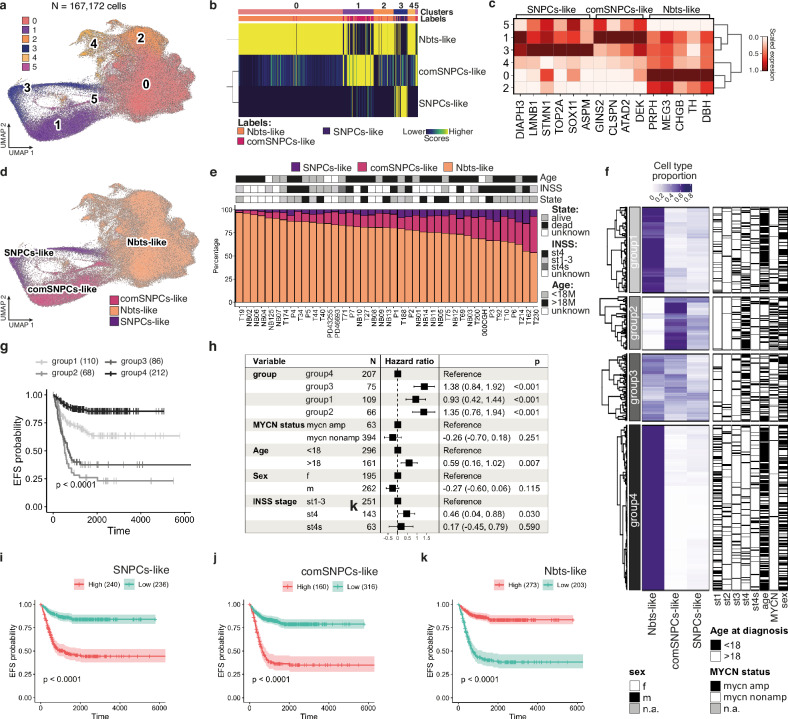
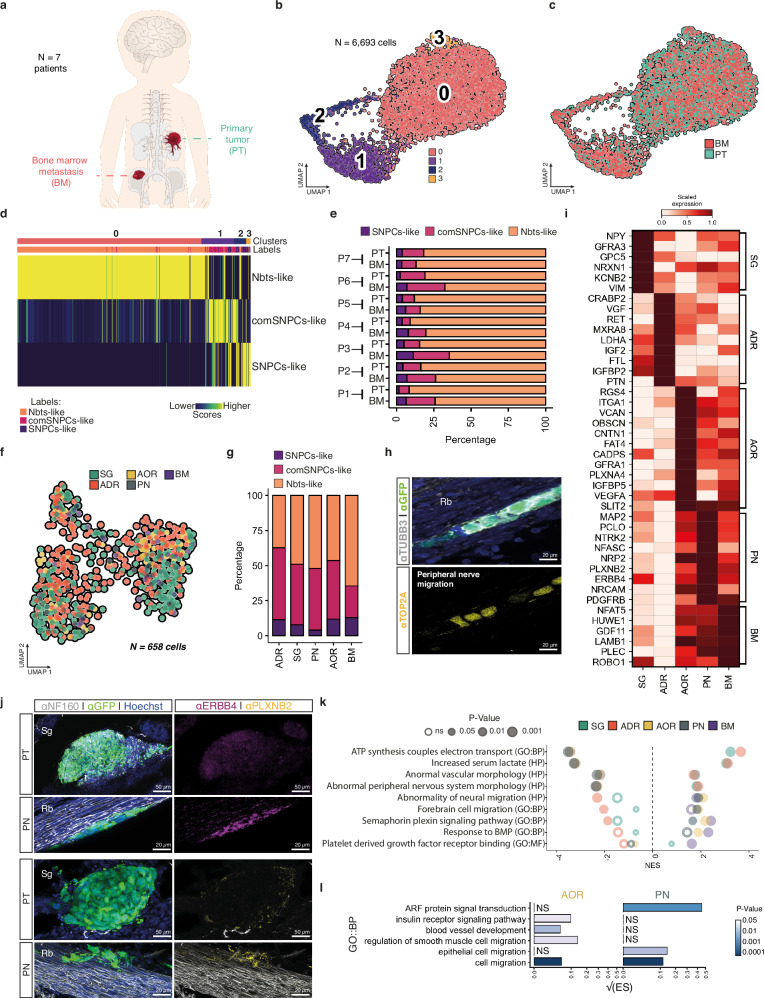
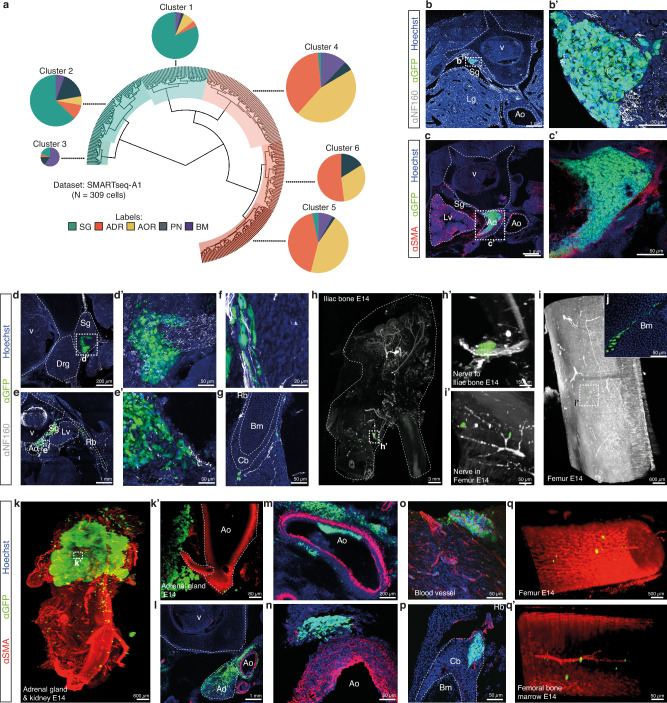
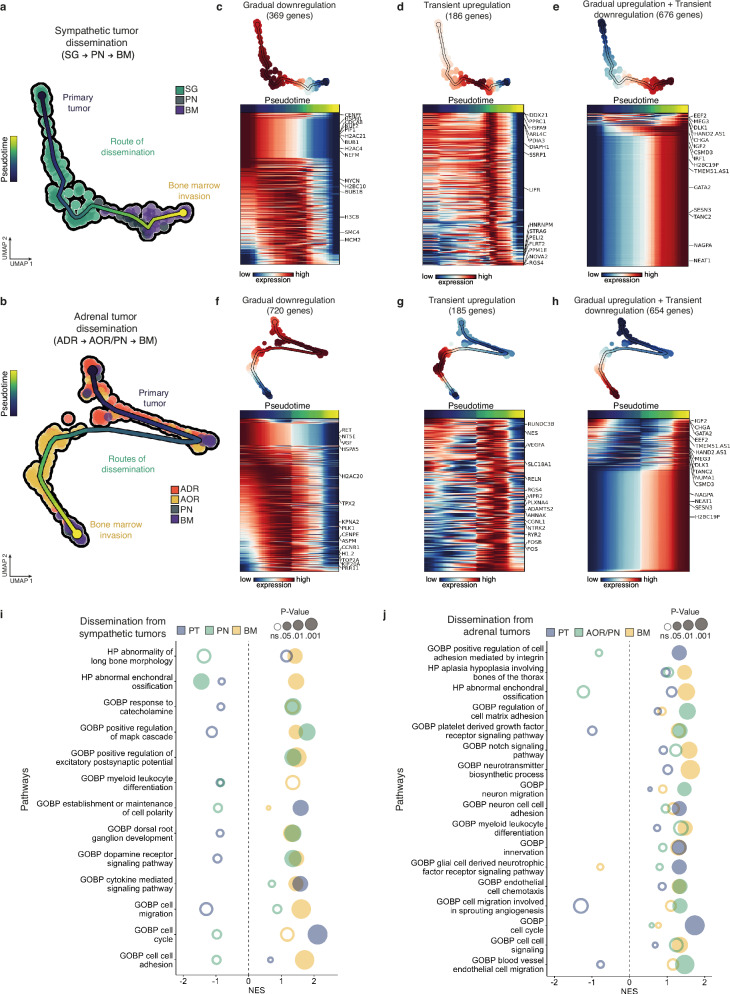
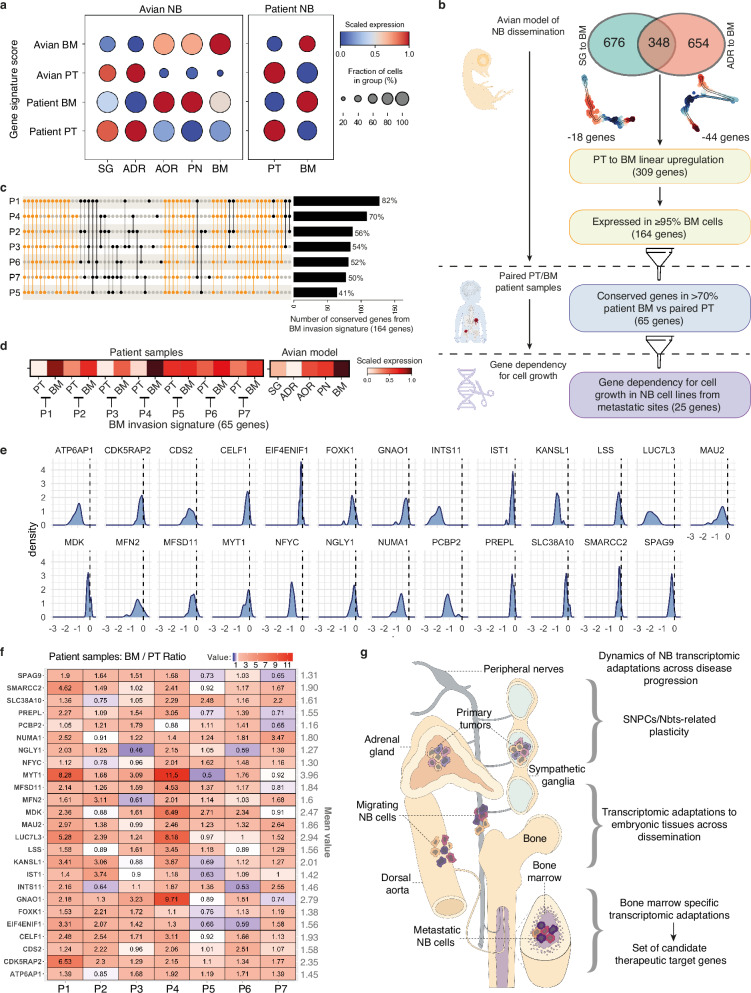
Despite their indisputable importance in neuroblastoma (NB) pathology, knowledge of the bases of NB plasticity and heterogeneity remains incomplete. They may be rooted in developmental trajectories of their lineage of origin, the sympatho-adrenal neural crest. We find that implanting human NB cells in the neural crest of the avian embryo allows recapitulating the metastatic sequence until bone marrow involvement. Using deep single cell RNA sequencing, we characterize transcriptome states of NB cells and their dynamics over time and space, and compare them to those of fetal sympatho-adrenal tissues and patient tumors and bone marrow samples. Here we report remarkable transcriptomic proximities restricted to an early sympathetic neuroblast branch that co-exist with phenotypical adaptations over disease progression and recapitulate intratumor and interpatient heterogeneity. Combining avian and patient datasets, we identify a list of genes upregulated during bone marrow involvement and associated with growth dependency, validating the relevance of our multimodal approach.
© 2024. The Author(s).
Conflict of interest statement
V.C. and C.D.B. declare the following competing interest: both are scientific advisors for the European Research Biological Center (ERBC) group. G.A. is employed by Cellenion, a BICO company. B.V., F.R., A.B., O.I., K.T., E.T., I.T., S.C., J.L., J.J.M., and G.T. declare no competing interests.
Figures
References
-
- Kattner, P. et al. Compare and contrast: pediatric cancer versus adult malignancies. Cancer Metastasis Rev.38, 673–682 (2019). - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
