

Enzyme-Responsive DNA Condensates
- PMID: 39503320
- PMCID: PMC11583213
- DOI: 10.1021/jacs.4c08919
Enzyme-Responsive DNA Condensates
Abstract
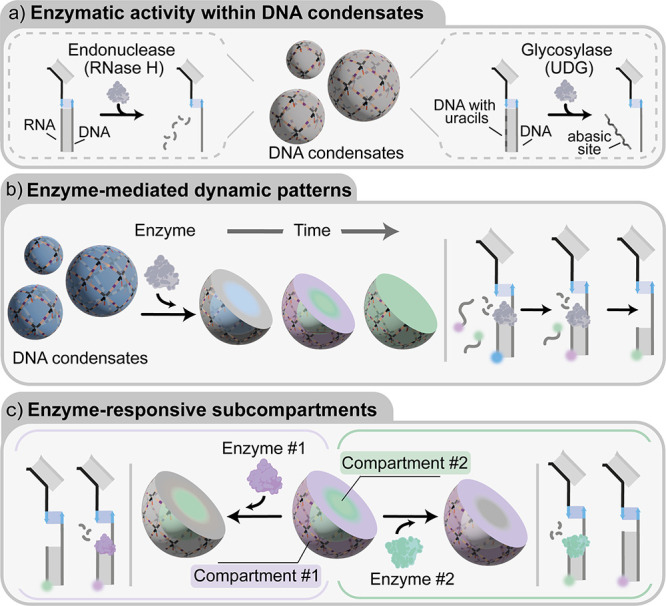
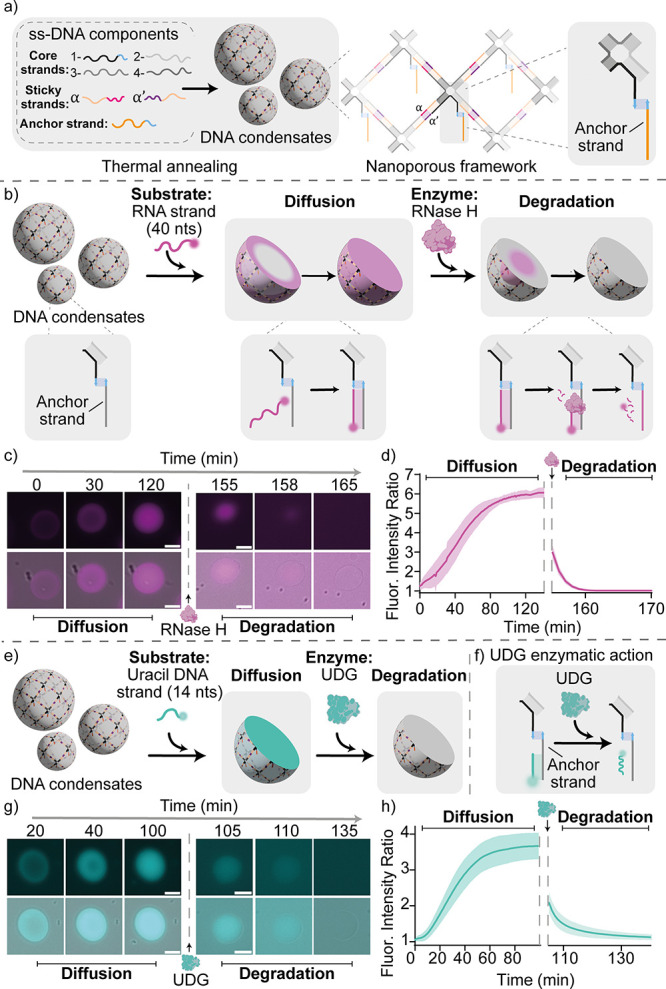
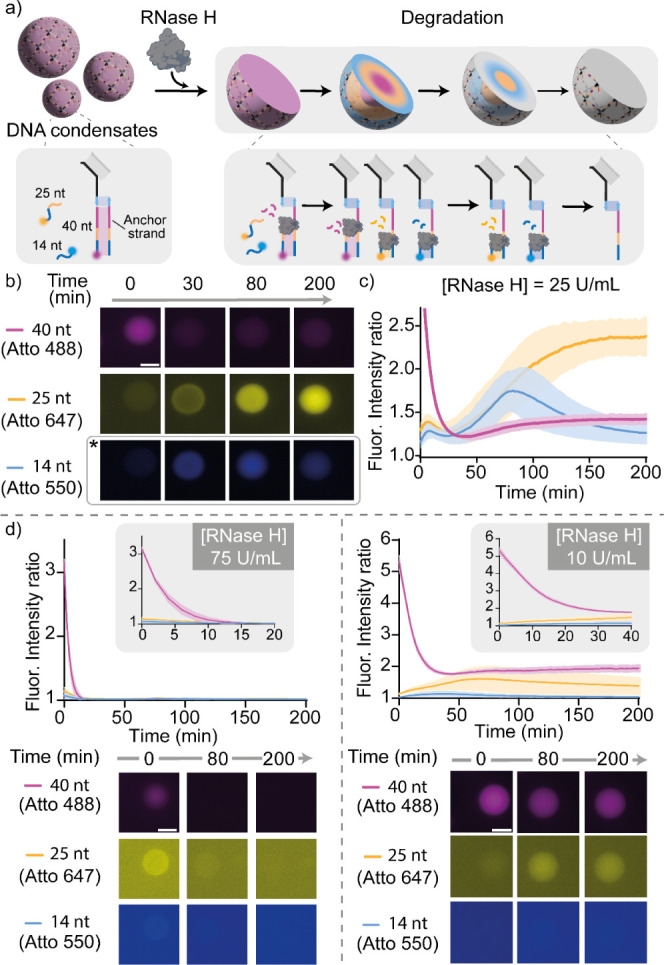
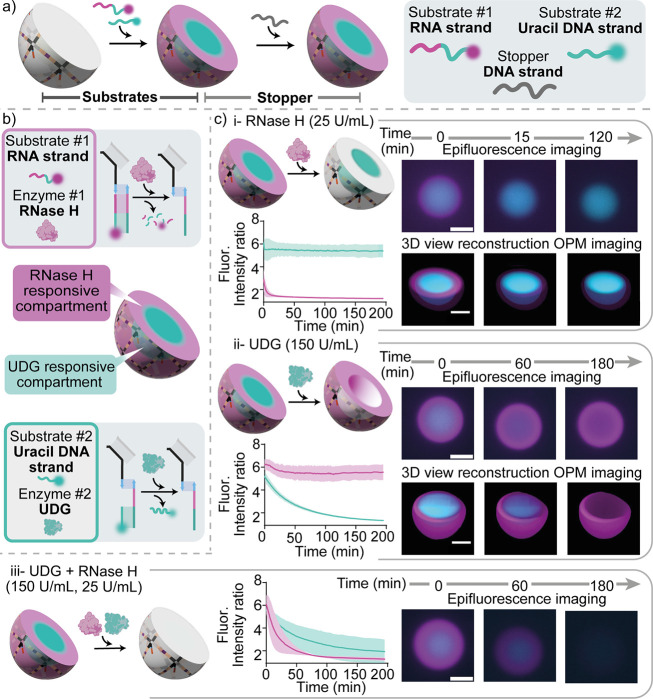
Membrane-less compartments and organelles are widely acknowledged for their role in regulating cellular processes, and there is an urgent need to harness their full potential as both structural and functional elements of synthetic cells. Despite rapid progress, synthetically recapitulating the nonequilibrium, spatially distributed responses of natural membrane-less organelles remains elusive. Here, we demonstrate that the activity of nucleic-acid cleaving enzymes can be localized within DNA-based membrane-less compartments by sequestering the respective DNA or RNA substrates. Reaction-diffusion processes lead to complex nonequilibrium patterns, dependent on enzyme concentration. By arresting similar dynamic patterns, we spatially organize different substrates in concentric subcompartments, which can be then selectively addressed by different enzymes, demonstrating spatial distribution of enzymatic activity. Besides expanding our ability to engineer advanced biomimetic functions in synthetic membrane-less organelles, our results may facilitate the deployment of DNA-based condensates as microbioreactors or platforms for the detection and quantitation of enzymes and nucleic acids.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
