

Enhanced biosynthesis of poly(3-hydroxybutyrate) in engineered strains of Pseudomonas putida via increased malonyl-CoA availability
- PMID: 39503721
- PMCID: PMC11539682
- DOI: 10.1111/1751-7915.70044
Enhanced biosynthesis of poly(3-hydroxybutyrate) in engineered strains of Pseudomonas putida via increased malonyl-CoA availability
Abstract
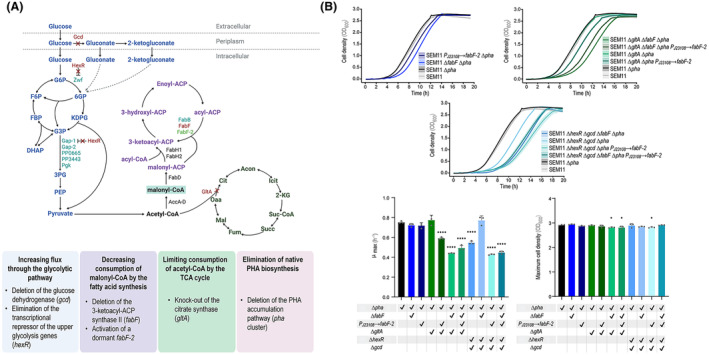
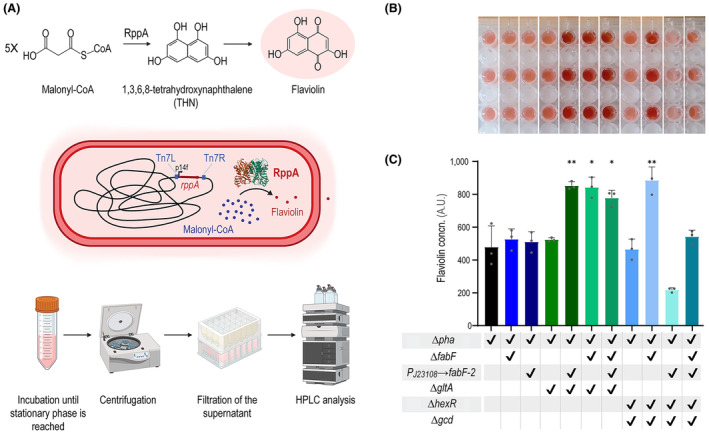
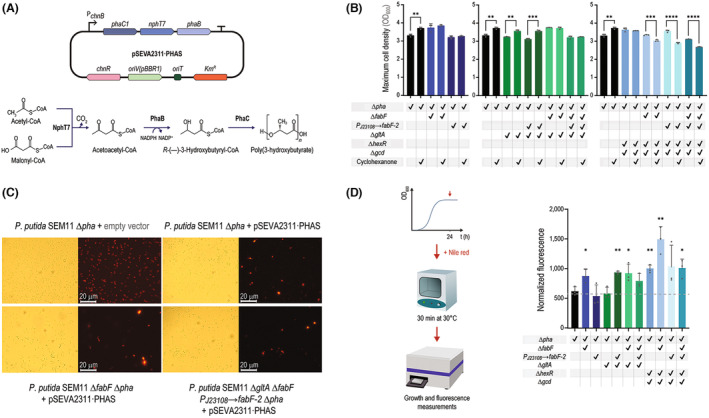
Malonyl-coenzyme A (CoA) is a key precursor for the biosynthesis of multiple value-added compounds by microbial cell factories, including polyketides, carboxylic acids, biofuels, and polyhydroxyalkanoates. Owing to its role as a metabolic hub, malonyl-CoA availability is limited by competition in several essential metabolic pathways. To address this limitation, we modified a genome-reduced Pseudomonas putida strain to increase acetyl-CoA carboxylation while limiting malonyl-CoA utilization. Genes involved in sugar catabolism and its regulation, the tricarboxylic acid (TCA) cycle, and fatty acid biosynthesis were knocked-out in specific combinations towards increasing the malonyl-CoA pool. An enzyme-coupled biosensor, based on the rppA gene, was employed to monitor malonyl-CoA levels in vivo. RppA is a type III polyketide synthase that converts malonyl-CoA into flaviolin, a red-colored polyketide. We isolated strains displaying enhanced malonyl-CoA availability via a colorimetric screening method based on the RppA-dependent red pigmentation; direct flaviolin quantification identified four engineered strains had a significant increase in malonyl-CoA levels. We further modified these strains by adding a non-canonical pathway that uses malonyl-CoA as precursor for poly(3-hydroxybutyrate) biosynthesis. These manipulations led to increased polymer accumulation in the fully engineered strains, validating our general strategy to boost the output of malonyl-CoA-dependent pathways in P. putida.
© 2024 The Author(s). Microbial Biotechnology published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Repurposing type III polyketide synthase as a malonyl-CoA biosensor for metabolic engineering in bacteria.Proc Natl Acad Sci U S A. 2018 Oct 2;115(40):9835-9844. doi: 10.1073/pnas.1808567115. Epub 2018 Sep 19. Proc Natl Acad Sci U S A. 2018. PMID: 30232266 Free PMC article.
-
Computer-assisted multilevel optimization of malonyl-CoA availability in Pseudomonas putida.Metab Eng. 2025 Jul;90:165-177. doi: 10.1016/j.ymben.2025.03.008. Epub 2025 Mar 17. Metab Eng. 2025. PMID: 40107409
-
A Pseudomonas taiwanensis malonyl-CoA platform strain for polyketide synthesis.Metab Eng. 2023 May;77:219-230. doi: 10.1016/j.ymben.2023.04.001. Epub 2023 Apr 8. Metab Eng. 2023. PMID: 37031949
-
Engineering intracellular malonyl-CoA availability in microbial hosts and its impact on polyketide and fatty acid synthesis.Appl Microbiol Biotechnol. 2020 Jul;104(14):6057-6065. doi: 10.1007/s00253-020-10643-7. Epub 2020 May 8. Appl Microbiol Biotechnol. 2020. PMID: 32385515 Free PMC article. Review.
-
Design and application of genetically-encoded malonyl-CoA biosensors for metabolic engineering of microbial cell factories.Metab Eng. 2017 Nov;44:253-264. doi: 10.1016/j.ymben.2017.10.011. Epub 2017 Oct 31. Metab Eng. 2017. PMID: 29097310 Review.
Cited by
-
Leveraging Engineered Pseudomonas putida Minicells for Bioconversion of Organic Acids into Short-Chain Methyl Ketones.ACS Synth Biol. 2025 Jan 17;14(1):257-272. doi: 10.1021/acssynbio.4c00700. Epub 2025 Jan 3. ACS Synth Biol. 2025. PMID: 39748701 Free PMC article.
-
Seven critical challenges in synthetic one-carbon assimilation and their potential solutions.FEMS Microbiol Rev. 2025 Jan 14;49:fuaf011. doi: 10.1093/femsre/fuaf011. FEMS Microbiol Rev. 2025. PMID: 40175298 Free PMC article. Review.
-
Improving Geldanamycin Production in Streptomyces geldanamycininus Through UV Mutagenesis of Protoplast.Microorganisms. 2025 Jan 17;13(1):186. doi: 10.3390/microorganisms13010186. Microorganisms. 2025. PMID: 39858954 Free PMC article.
References
-
- Ackermann, Y.S. , de Witt, J. , Mezzina, M.P. , Schroth, C. , Polen, T. , Nikel, P.I. et al. (2024) Bio‐upcycling of even and uneven medium‐chain‐length diols and dicarboxylates to polyhydroxyalkanoates using engineered Pseudomonas putida . Microbial Cell Factories, 23, 54. Available from: 10.1186/s12934-024-02310-7 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
