

GLIS3: A novel transcriptional regulator of mitochondrial functions and metabolic reprogramming in postnatal kidney and polycystic kidney disease
- PMID: 39505148
- PMCID: PMC11613186
- DOI: 10.1016/j.molmet.2024.102052
GLIS3: A novel transcriptional regulator of mitochondrial functions and metabolic reprogramming in postnatal kidney and polycystic kidney disease
Abstract
Objectives: Deficiency in the transcription factor (TF) GLI-Similar 3 (GLIS3) in humans and mice leads to the development of polycystic kidney disease (PKD). In this study, we investigate the role of GLIS3 in the regulation of energy metabolism and mitochondrial functions in relation to its role in normal kidney and metabolic reprogramming in PKD pathogenesis.
Methods: Transcriptomics, cistromics, and metabolomics were used to obtain insights into the role of GLIS3 in the regulation of energy homeostasis and mitochondrial metabolism in normal kidney and PKD pathogenesis using GLIS3-deficient mice.
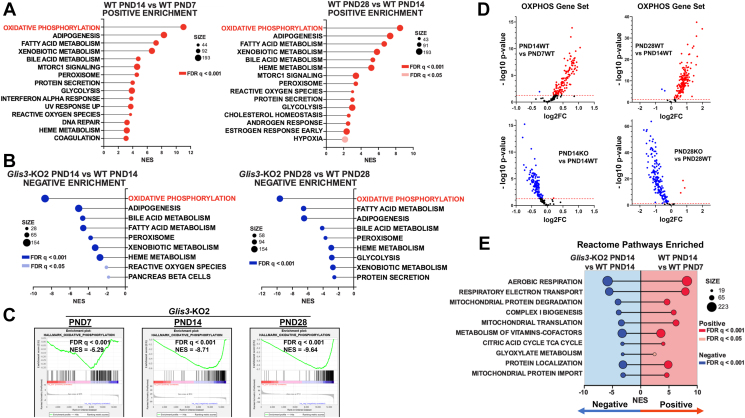
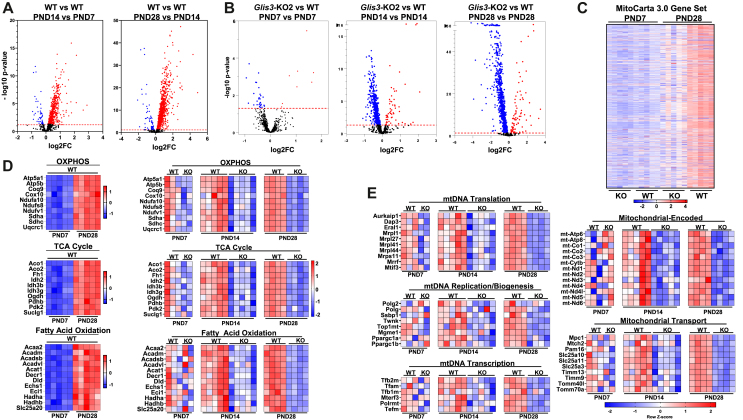
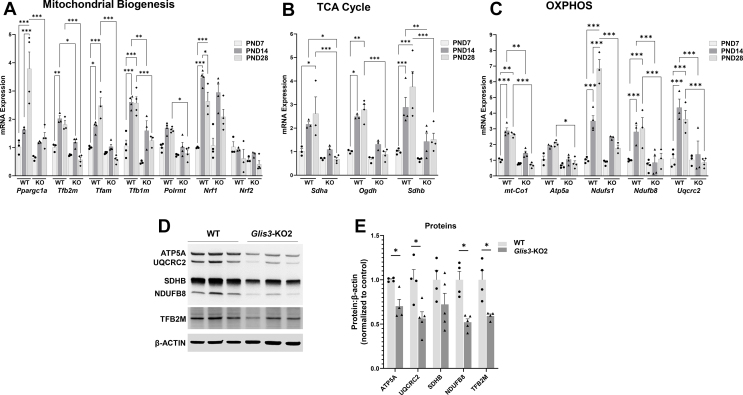
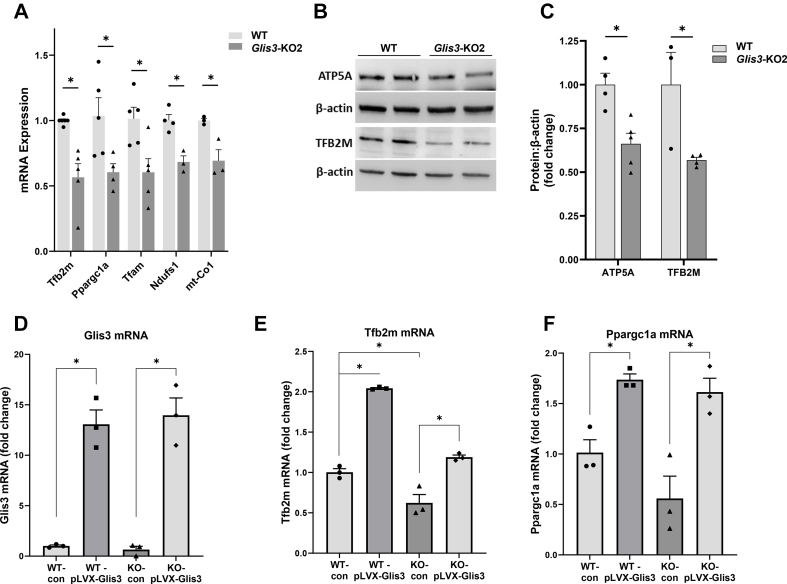
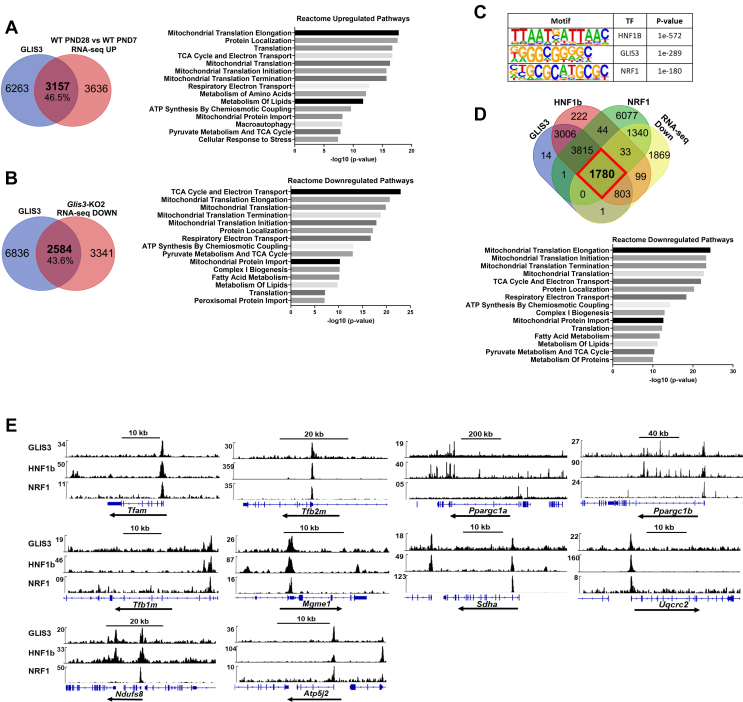
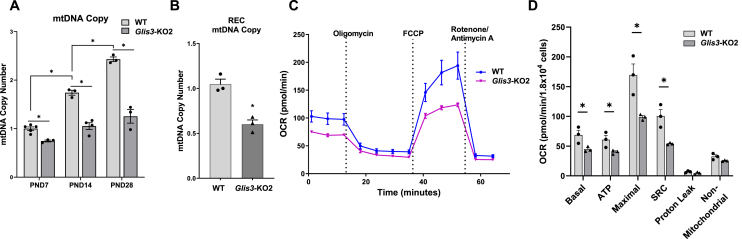
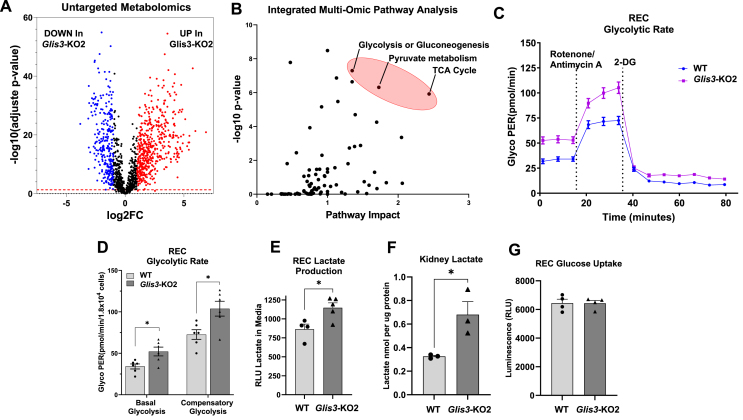
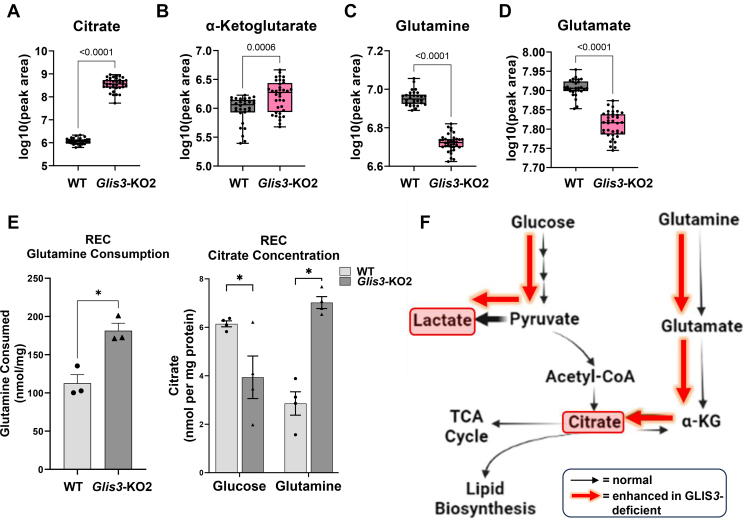
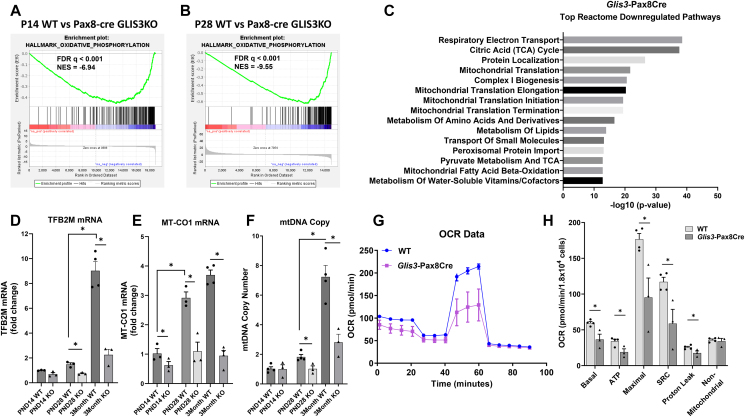
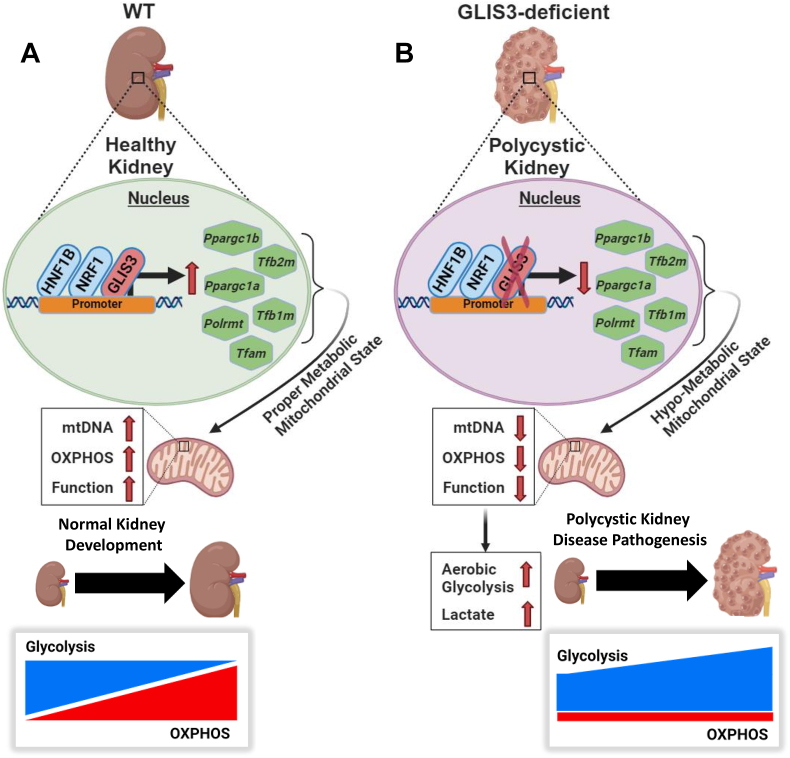
Results: Transcriptome analysis showed that many genes critical for mitochondrial biogenesis, oxidative phosphorylation (OXPHOS), fatty acid oxidation (FAO), and the tricarboxylic acid (TCA) cycle, including Tfam, Tfb1m, Tfb2m, Ppargc1a, Ppargc1b, Atp5j2, Hadha, and Sdha, are significantly suppressed in kidneys from both ubiquitous and tissue-specific Glis3-deficient mice. ChIP-Seq analysis demonstrated that GLIS3 is associated with the regulatory region of many of these genes, indicating that their transcription is directly regulated by GLIS3. Cistrome analyses revealed that GLIS3 binding loci frequently located near those of hepatocyte nuclear factor 1-Beta (HNF1B) and nuclear respiratory factor 1 (NRF1) suggesting GLIS3 regulates transcription of many metabolic and mitochondrial function-related genes in coordination with these TFs. Seahorse analysis and untargeted metabolomics corroborated that mitochondrial OXPHOS utilization is suppressed in GLIS3-deficient kidneys and showed that key metabolites in glycolysis, TCA cycle, and glutamine pathways were altered indicating increased reliance on aerobic glycolysis and glutamine anaplerosis. These features of metabolic reprogramming may contribute to a bioenergetic environment that supports renal cyst formation and progression in Glis3-deficient mice kidneys.
Conclusions: We identify GLIS3 as a novel positive regulator of the transition from aerobic glycolysis to OXPHOS in normal early postnatal kidney development by directly regulating the transcription of mitochondrial metabolic genes. Loss of GLIS3 induces several features of renal cell metabolic reprogramming. Our study identifies GLIS3 as a new participant in an interconnected transcription regulatory network, that includes HNF1B and NRF1, critical in the regulation of mitochondrial-related gene expression and energy metabolism in normal postnatal kidneys and PKD pathogenesis in Glis3-deficient mice.
Keywords: Aerobic glycolysis; GLIS3; Metabolic reprogramming; Oxidative phosphorylation; Polycystic kidney disease; Transcription.
Published by Elsevier GmbH.
Conflict of interest statement
Declaration of competing interest All the authors declared no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
