

Substrate specificity and kinetic mechanism of 3β-hydroxy-Δ5-C27-steroid oxidoreductase
- PMID: 39505210
- PMCID: PMC11648244
- DOI: 10.1016/j.jbc.2024.107945
Substrate specificity and kinetic mechanism of 3β-hydroxy-Δ5-C27-steroid oxidoreductase
Abstract
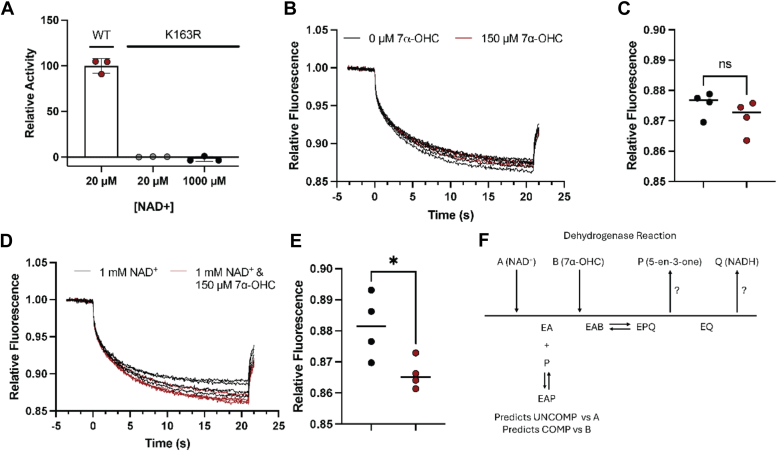
Cholesterol is a key sterol whose homeostasis is primarily maintained through bile acid metabolism. Proper bile acid formation is vital for nutrient and fat-soluble vitamin absorption and emulsification of lipids. Synthesis of bile acids occurs through two main pathways, both of which rely on 3β-hydroxy-Δ5-C27-steroid oxidoreductase (HSD3B7) to begin epimerization of the 3β hydroxyl of cholesterol into its active 3α conformation. In this sequence, HSD3B7 catalyzes the dehydrogenation of the 3β-hydroxy group followed by isomerization of the Δ5-cholestene-3-one. These reactions are some of the many steps that transform cholesterol for either storage or secretion. HSD3B7 has distinct activity from other 3β-HSD family members leaving significant gaps in our understanding of its mode of catalysis and substrate specificity. In addition, the role of HSD3B7 in health and disease positions it as a metabolic vulnerability that could be harnessed as a therapeutic target. To this end, we evaluated the mechanism of HSD3B7 catalysis and reveal that HSD3B7 displays activity toward diverse 7α-hydroxylated oxysterols. HSD3B7 retains its catalytic efficiency toward these substrates, suggesting that its substrate binding pocket can withstand changes in polarity upon alterations to this hydrocarbon tail. Experiments aimed at determining substrate order are consistent with HSD3B7 catalyzing a sequential ordered bi-bi reaction mechanism with the binding of NAD+ followed by 7α-hydroxycholesterol to form a central complex. HSD3B7 bifunctional activity is dependent on membrane localization through a putative membrane-associated helix giving insight into potential regulation of enzyme activity. We found strong binding of the NADH product thought to activate the isomerization reaction. Homology models of HSD3B7 reveal a potential substrate pocket that allows for oxysterol binding, and mutagenesis was utilized to support this model. Together, these studies offer an understanding of substrate specificity and kinetic mechanism of HSD3B7, which can be exploited for future drug development.
Keywords: bile acid synthesis; cholesterol metabolism; enzyme mechanism; membrane protein; metabolism; oxidoreductase.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article except for T.M.P. who is the cofounder of Penzymes and is a consultant for Sage Therapeutics and Propella, Inc.
Figures






References
-
- Holick M.F.F., J E., McNeill S.C., Richtand N.M., Henley J.W., Potts J.T. Photometabolism of 7-hydroxycholesterol to previtamin D3 in skin. Biochem. Biophysical Res. Commun. 1977;76:107–114. - PubMed
-
- Javitt N.B. Bile acid synthesis from cholesterol: regulatory and auxillary pathways. FASEB J. 1994;8:1308–1311. - PubMed
-
- Payne A.H., Hales D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004;25:947–970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
