

Induction of a distinct macrophage population and protection from lung injury and fibrosis by Notch2 blockade
- PMID: 39505846
- PMCID: PMC11541919
- DOI: 10.1038/s41467-024-53700-9
Induction of a distinct macrophage population and protection from lung injury and fibrosis by Notch2 blockade
Abstract
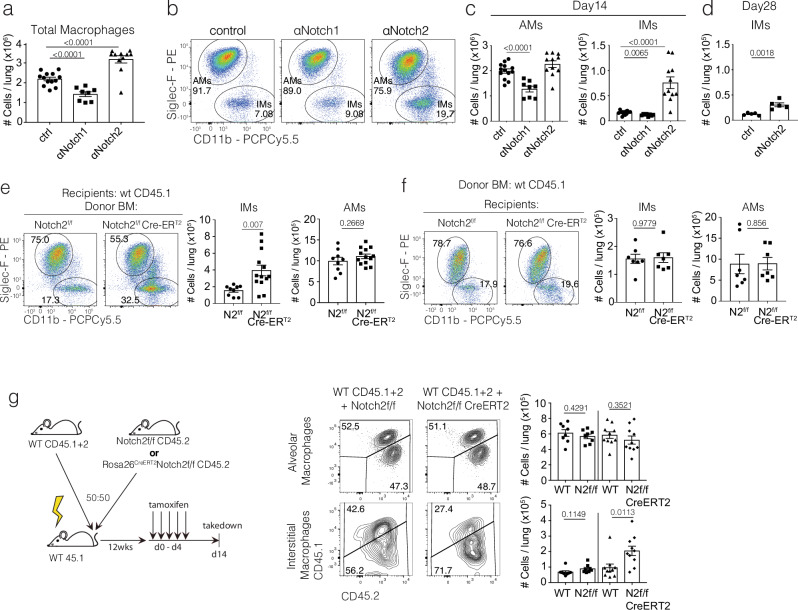
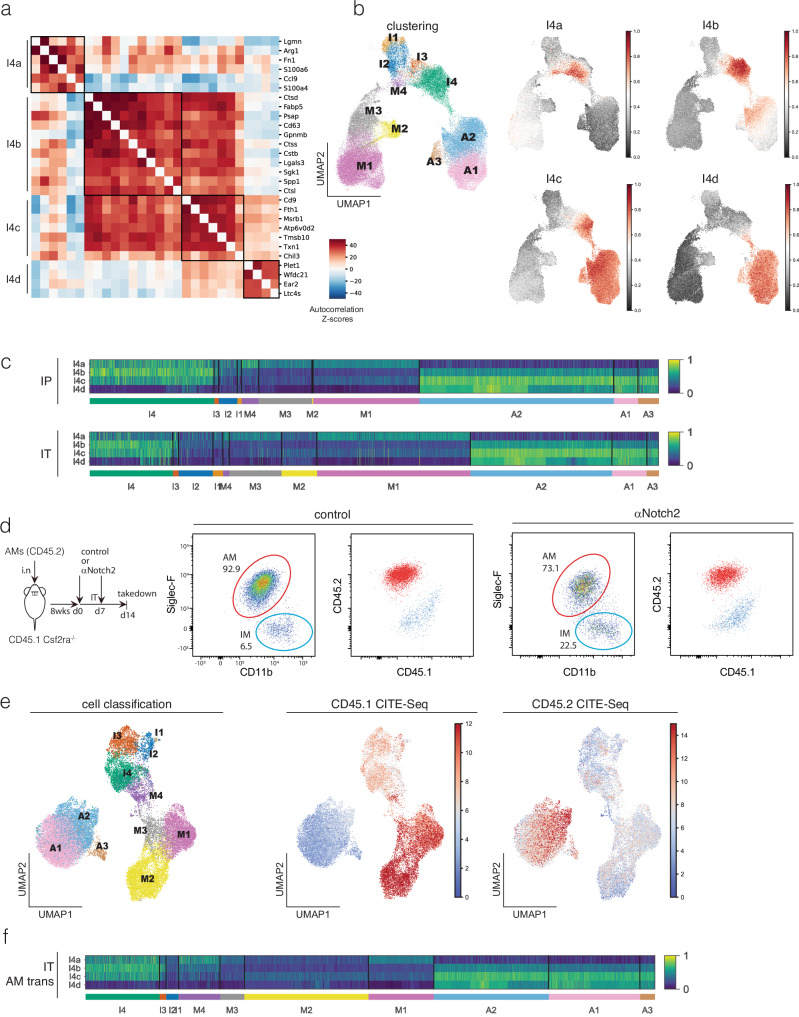
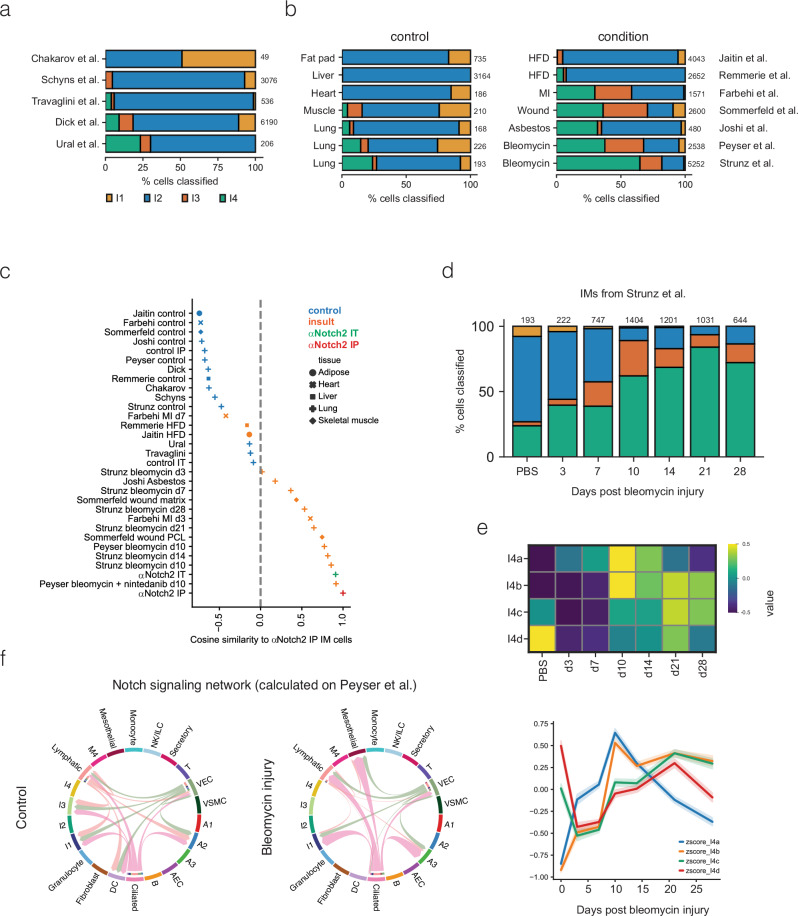
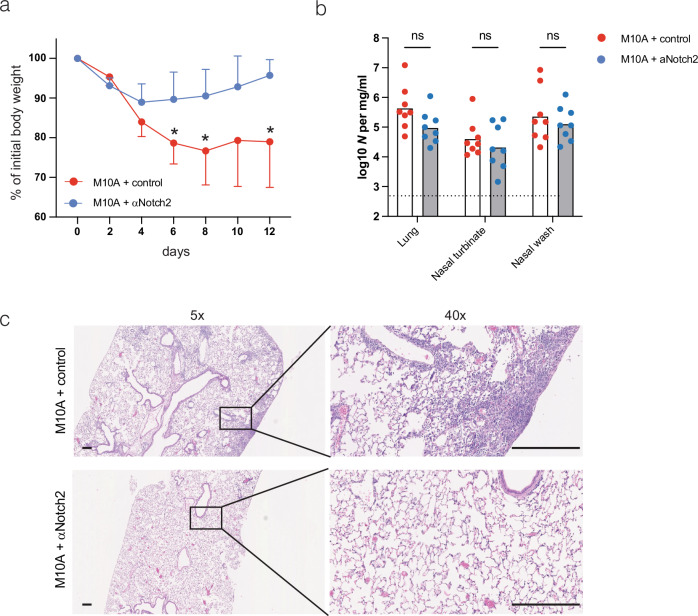
Macrophages are pleiotropic and diverse cells that populate all tissues of the body. Besides tissue-specific resident macrophages such as alveolar macrophages, Kupffer cells, and microglia, multiple organs harbor at least two subtypes of other resident macrophages at steady state. During certain circumstances, like tissue insult, additional subtypes of macrophages are recruited to the tissue from the monocyte pool. Previously, a recruited macrophage population marked by expression of Spp1, Cd9, Gpnmb, Fabp5, and Trem2, has been described in several models of organ injury and cancer, and has been linked to fibrosis in mice and humans. Here, we show that Notch2 blockade, given systemically or locally, leads to an increase in this putative pro-fibrotic macrophage in the lung and that this macrophage state can only be adopted by monocytically derived cells and not resident alveolar macrophages. Using a bleomycin and COVID-19 model of lung injury and fibrosis, we find that the expansion of these macrophages before lung injury does not promote fibrosis but rather appears to ameliorate it. This suggests that these damage-associated macrophages are not, by themselves, drivers of fibrosis in the lung.
© 2024. The Author(s).
Conflict of interest statement
M.C.T., S.P.M., A.W., A.A., J.D.W., J.La, J.J.H., V.S.G., M. De S. B.K., S.A., J. Lu, S.J., A.R., H.B., C.D.A., S.Da, Z.M., H.B., S.Du, A.S.S., and M.N. are employees of Genentech/Roche. The remaining authors declare no other competing interests.
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
