

Repeat mediated excision of gene drive elements for restoring wild-type populations
- PMID: 39509462
- PMCID: PMC11584131
- DOI: 10.1371/journal.pgen.1011450
Repeat mediated excision of gene drive elements for restoring wild-type populations
Abstract
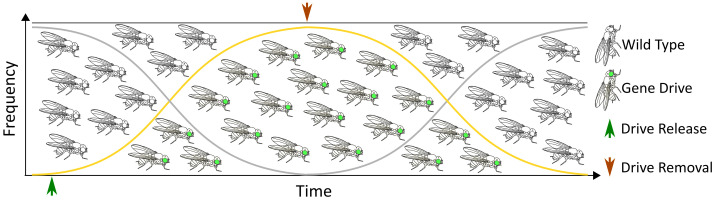
Here, we demonstrate that single strand annealing (SSA) can be co-opted for the precise autocatalytic excision of a drive element. We have termed this technology Repeat Mediated Excision of a Drive Element (ReMEDE). By engineering direct repeats flanking the drive allele and inducing a double-strand DNA break (DSB) at a second endonuclease target site within the allele, we increased the utilization of SSA repair. ReMEDE was incorporated into the mutagenic chain reaction (MCR) gene drive targeting the yellow gene of Drosophila melanogaster, successfully replacing drive alleles with wild-type alleles. Sequencing across the Cas9 target site confirmed transgene excision by SSA after pair-mated outcrosses with yReMEDE females, revealing ~4% inheritance of an engineered silent TcG marker sequence. However, phenotypically wild-type flies with alleles of indeterminate biogenesis also were observed, retaining the TGG sequence (~16%) or harboring a silent gGG mutation (~0.5%) at the PAM site. Additionally, ~14% of alleles in the F2 flies were intact or uncut paternally inherited alleles, indicating limited maternal deposition of Cas9 RNP. Although ReMEDE requires further research and development, the technology has some promising features as a gene drive mitigation strategy, notably its potential to restore wild-type populations without additional transgenic releases or large-scale environmental modifications.
Copyright: © 2024 Chennuri et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: KMM and ZNA are inventors on US provisional patent application PCT/US2021/041951, submitted by Texas A&M University, which covers vector constructs that are pre-programmed to self-eliminate or self- remove at a predetermined time, and methods of making the same. PRC, JZ, RDM, MLNM declare no competing interests.
Figures





Update of
-
Repeat mediated excision of gene drive elements for restoring wild-type populations.bioRxiv [Preprint]. 2023 Nov 23:2023.11.23.568397. doi: 10.1101/2023.11.23.568397. bioRxiv. 2023. Update in: PLoS Genet. 2024 Nov 7;20(11):e1011450. doi: 10.1371/journal.pgen.1011450. PMID: 38045402 Free PMC article. Updated. Preprint.
Similar articles
-
Repeat mediated excision of gene drive elements for restoring wild-type populations.bioRxiv [Preprint]. 2023 Nov 23:2023.11.23.568397. doi: 10.1101/2023.11.23.568397. bioRxiv. 2023. Update in: PLoS Genet. 2024 Nov 7;20(11):e1011450. doi: 10.1371/journal.pgen.1011450. PMID: 38045402 Free PMC article. Updated. Preprint.
-
A self-eliminating allelic-drive reverses insecticide resistance in Drosophila leaving no transgene in the population.Nat Commun. 2024 Nov 17;15(1):9961. doi: 10.1038/s41467-024-54210-4. Nat Commun. 2024. PMID: 39551783 Free PMC article.
-
Inherently confinable split-drive systems in Drosophila.Nat Commun. 2021 Mar 5;12(1):1480. doi: 10.1038/s41467-021-21771-7. Nat Commun. 2021. PMID: 33674604 Free PMC article.
-
The dawn of active genetics.Bioessays. 2016 Jan;38(1):50-63. doi: 10.1002/bies.201500102. Epub 2015 Dec 10. Bioessays. 2016. PMID: 26660392 Free PMC article. Review.
-
Single-strand annealing: Molecular mechanisms and potential applications in CRISPR-Cas-based precision genome editing.Biotechnol J. 2022 Jul;17(7):e2100413. doi: 10.1002/biot.202100413. Epub 2021 Dec 10. Biotechnol J. 2022. PMID: 34846104 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
