

Structure-function analysis of tRNA t6A-catalysis, assembly, and thermostability of Aquifex aeolicus TsaD2B2 tetramer in complex with TsaE
- PMID: 39510188
- PMCID: PMC11648230
- DOI: 10.1016/j.jbc.2024.107962
Structure-function analysis of tRNA t6A-catalysis, assembly, and thermostability of Aquifex aeolicus TsaD2B2 tetramer in complex with TsaE
Abstract
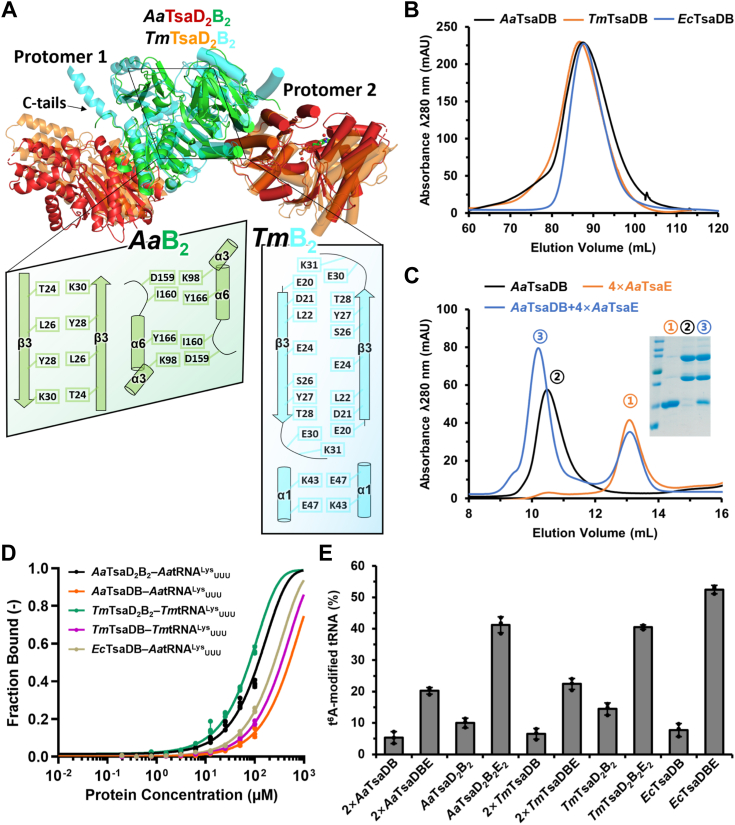
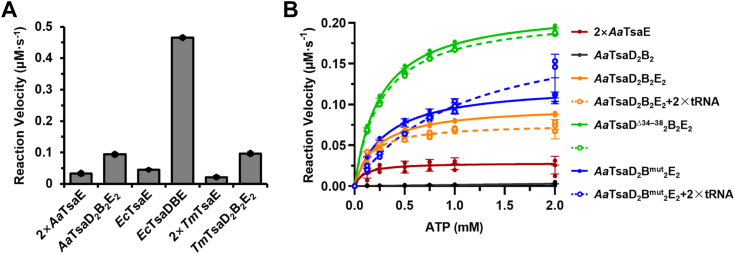
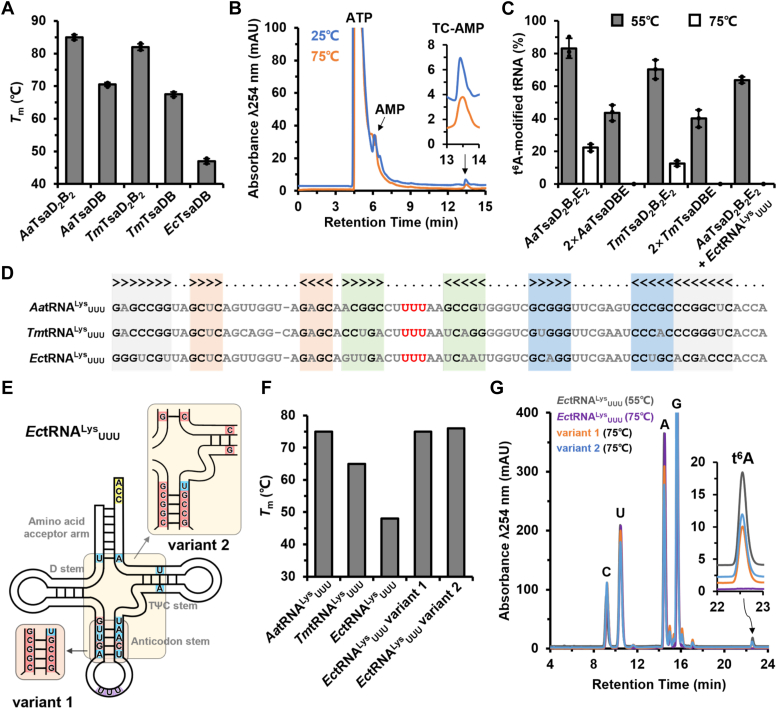
The universal N6-threonylcarbamoyladenosine (t6A) at position 37 of tRNAs is one of the core post-transcriptional modifications that are needed for promoting translational fidelity. In bacteria, TsaC uses L-threonine, bicarbonate, and ATP to generate an intermediate threonylcarbamoyladenylate (TC-AMP), of which the TC moiety is transferred to N6 atom of tRNA A37 to generate t6A by TsaD with the support of TsaB and TsaE. TsaD and TsaB form a TsaDB dimer to which tRNA and TsaE are competitively bound. The catalytic mechanism of TsaD and auxiliary roles of TsaB and TsaE remain to be fully elucidated. In this study, we reconstituted tRNA t6A biosynthesis using TsaC, TsaD, TsaB, and TsaE from Aquifex aeolicus and determined crystal structures of apo-form and ADP-bound form of TsaD2B2 tetramer. Our TsaD2B2-TsaE-tRNA model coupled with functional validations reveal that the binding of tRNA or TsaE to TsaDB is regulated by C-terminal tail of TsaB and a helical hairpin α1-α2 of TsaD. A. aeolicus TsaDB possesses a basal t6A catalytic activity that is stimulated by TsaE at the cost of ATP consumption. Our data suggest that the binding of TsaE to TsaDB induces conformational changes of α1, α2, α6, α7, and α8 of TsaD and C-terminal tail of TsaB, leading to the release of tRNA t6A and AMP. ATP-mediated binding of TsaE to TsaDB resets a t6A active conformation of TsaDB. Dimerization of TsaDB enhances thermostability and promotes t6A catalysis of TsaD2B2-tRNA, of which GC base pairs in anticodon stem are needed for the correct folding of thermophilic tRNA at higher temperatures.
Keywords: ATP hydrolysis; Aquifex aeolicus; GC base pairs content; TsaD(2)B(2) tetramer; TsaD–TsaB–TsaE–tRNA assembly; crystal structure; enzymatic reconstitution; oligomerization; tRNA t(6)A; thermostability.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures


References
-
- Grosjean H., Sprinzl M., Steinberg S. Posttranscriptionally modified nucleosides in transfer RNA: their locations and frequencies. Biochimie. 1995;77:139–141. - PubMed
-
- Giege R., Juhling F., Putz J., Stadler P., Sauter C., Florentz C. Structure of transfer RNAs: similarity and variability. Wiley Interdiscip. Rev. RNA. 2012;3:37–61. - PubMed
-
- Bjork G.R., Ericson J.U., Gustafsson C.E., Hagervall T.G., Jonsson Y.H., Wikstrom P.M. Transfer RNA modification. Annu. Rev. Biochem. 1987;56:263–287. - PubMed
-
- Yarian C., Townsend H., Czestkowski W., Sochacka E., Malkiewicz A.J., Guenther R., et al. Accurate translation of the genetic code depends on tRNA modified nucleosides. J. Biol. Chem. 2002;277:16391–16395. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Miscellaneous
