

Cryo-EM structures of the membrane repair protein dysferlin
- PMID: 39511170
- PMCID: PMC11544258
- DOI: 10.1038/s41467-024-53773-6
Cryo-EM structures of the membrane repair protein dysferlin
Abstract
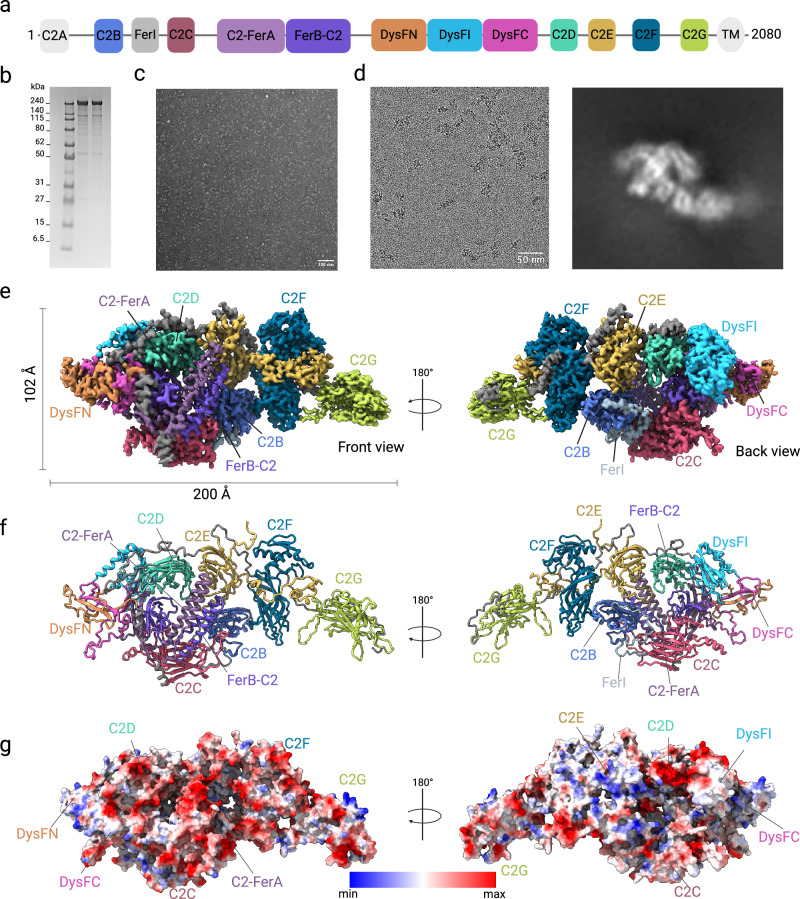
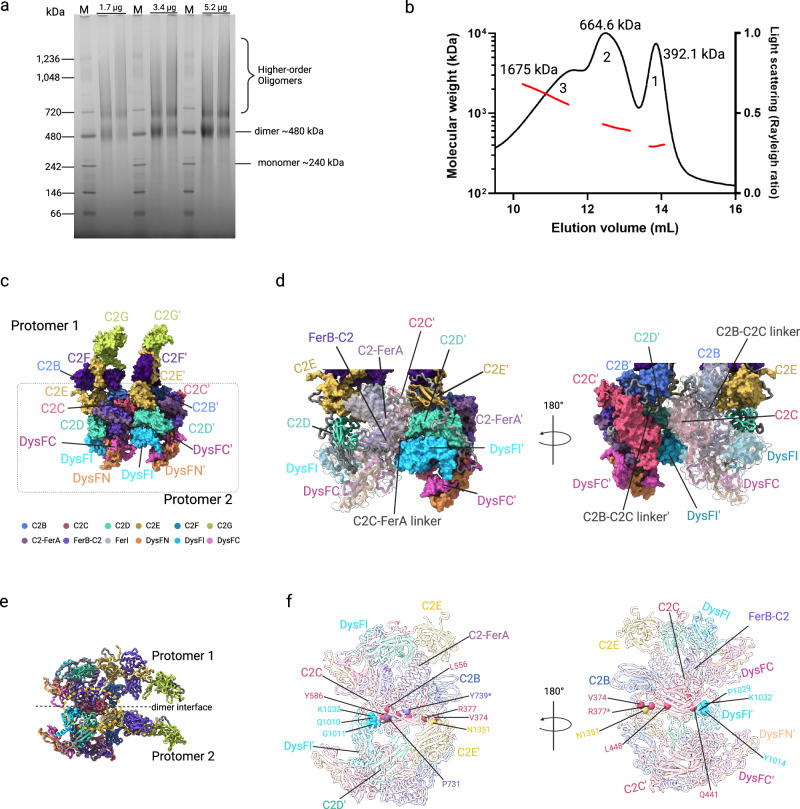
Plasma membrane repair in response to damage is essential for cell viability. The ferlin family protein dysferlin plays a key role in Ca2+-dependent membrane repair in striated muscles. Mutations in dysferlin lead to a spectrum of diseases known as dysferlinopathies. The lack of a structure of dysferlin and other ferlin family members has impeded a mechanistic understanding of membrane repair mechanisms and the development of therapies. Here, we present the cryo-EM structures of the full-length human dysferlin monomer and homodimer at 2.96 Å and 4.65 Å resolution. These structures define the architecture of dysferlin, ferlin family-specific domains, and homodimerization mechanisms essential to function. Furthermore, biophysical and cell biology studies revealed how missense mutations in dysferlin contribute to disease mechanisms. In summary, our study provides a framework for the molecular mechanisms of dysferlin and the broader ferlin family, offering a foundation for the development of therapeutic strategies aimed at treating dysferlinopathies.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Mutation impact on dysferlin inferred from database analysis and computer-based structural predictions.J Neurol Sci. 2006 Dec 1;250(1-2):71-8. doi: 10.1016/j.jns.2006.07.004. Epub 2006 Sep 22. J Neurol Sci. 2006. PMID: 16996541
-
FerA is a Membrane-Associating Four-Helix Bundle Domain in the Ferlin Family of Membrane-Fusion Proteins.Sci Rep. 2018 Jul 19;8(1):10949. doi: 10.1038/s41598-018-29184-1. Sci Rep. 2018. PMID: 30026467 Free PMC article.
-
Dysferlin function in skeletal muscle: Possible pathological mechanisms and therapeutical targets in dysferlinopathies.Exp Neurol. 2016 Sep;283(Pt A):246-54. doi: 10.1016/j.expneurol.2016.06.026. Epub 2016 Jun 25. Exp Neurol. 2016. PMID: 27349407 Review.
-
AMPK Complex Activation Promotes Sarcolemmal Repair in Dysferlinopathy.Mol Ther. 2020 Apr 8;28(4):1133-1153. doi: 10.1016/j.ymthe.2020.02.006. Epub 2020 Feb 12. Mol Ther. 2020. PMID: 32087766 Free PMC article.
-
Dysferlinopathies: Clinical and genetic variability.Clin Genet. 2022 Dec;102(6):465-473. doi: 10.1111/cge.14216. Epub 2022 Sep 6. Clin Genet. 2022. PMID: 36029111 Review.
Cited by
-
Whole Exome Sequencing Identified a Stop-Gained Mutation in DYSF Gene Associated With Dysferlinopathy in an Iranian Family.Int J Genomics. 2025 Jul 23;2025:9103068. doi: 10.1155/ijog/9103068. eCollection 2025. Int J Genomics. 2025. PMID: 40740503 Free PMC article.
-
Antisense oligonucleotide-mediated exon 27 skipping restores dysferlin function in dysferlinopathy patient-derived muscle cells.Mol Ther Nucleic Acids. 2025 Feb 4;36(1):102462. doi: 10.1016/j.omtn.2025.102462. eCollection 2025 Mar 11. Mol Ther Nucleic Acids. 2025. PMID: 39991471 Free PMC article. No abstract available.
-
Structural insights into lipid membrane binding by human ferlins.EMBO J. 2025 Jul;44(14):3926-3958. doi: 10.1038/s44318-025-00463-8. Epub 2025 May 28. EMBO J. 2025. PMID: 40437073 Free PMC article.
References
-
- Clarke, M. S., Caldwell, R. W., Chiao, H., Miyake, K. & McNeil, P. L. Contraction-induced cell wounding and release of fibroblast growth factor in heart. Circ. Res.76, 927–934 (1995). - PubMed
-
- McNeil, P. L. & Steinhardt, R. A. Plasma membrane disruption: repair, prevention, adaptation. Annu. Rev. Cell Dev. Biol.19, 697–731 (2003). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
