

The BAF complex enhances transcription through interaction with H3K56ac in the histone globular domain
- PMID: 39511190
- PMCID: PMC11544104
- DOI: 10.1038/s41467-024-53981-0
The BAF complex enhances transcription through interaction with H3K56ac in the histone globular domain
Abstract
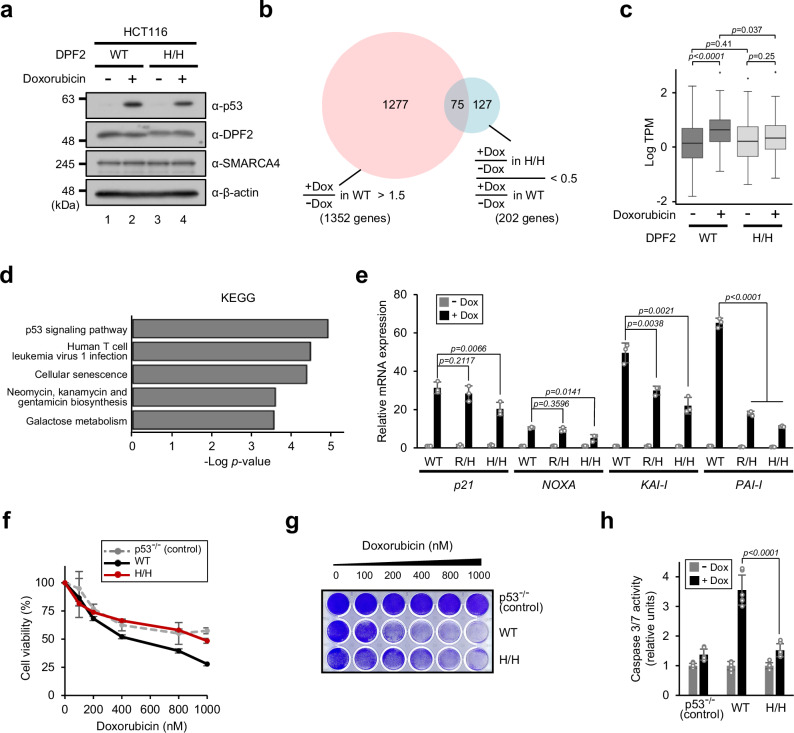
Histone post-translational modifications play pivotal roles in eukaryotic gene expression. To date, most studies have focused on modifications in unstructured histone N-terminal tail domains and their binding proteins. However, transcriptional regulation by chromatin-effector proteins that directly recognize modifications in histone globular domains has yet to be clearly demonstrated, despite the richness of their multiple modifications. Here, we show that the ATP-dependent chromatin-remodeling BAF complex stimulates p53-dependent transcription through direct interaction with H3K56ac located on the lateral surface of the histone globular domain. Mechanistically, the BAF complex recognizes nucleosomal H3K56ac via the DPF domain in the DPF2 subunit and exhibits enhanced nucleosome-remodeling activity in the presence of H3K56ac. We further demonstrate that a defect in H3K56ac-BAF complex interaction leads to impaired p53-dependent gene expression and DNA damage responses. Our study provides direct evidence that histone globular domain modifications participate in the regulation of gene expression.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Millán-Zambrano, G., Burton, A., Bannister, A. J. & Schneider, R. Histone post-translational modifications - cause and consequence of genome function. Nat. Rev. Genet.23, 563–580 (2022). - PubMed
-
- Mohammad, H. P., Barbash, O. & Creasy, C. L. Targeting epigenetic modifications in cancer therapy: erasing the roadmap to cancer. Nat. Med.25, 403–418 (2019). - PubMed
-
- Janssen, S. M. & Lorincz, M. C. Interplay between chromatin marks in development and disease. Nat. Rev. Genet.23, 137–153 (2022). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
