

FABP4-mediated lipid accumulation and lipolysis in tumor-associated macrophages promote breast cancer metastasis
- PMID: 39513934
- PMCID: PMC11548877
- DOI: 10.7554/eLife.101221
FABP4-mediated lipid accumulation and lipolysis in tumor-associated macrophages promote breast cancer metastasis
Abstract
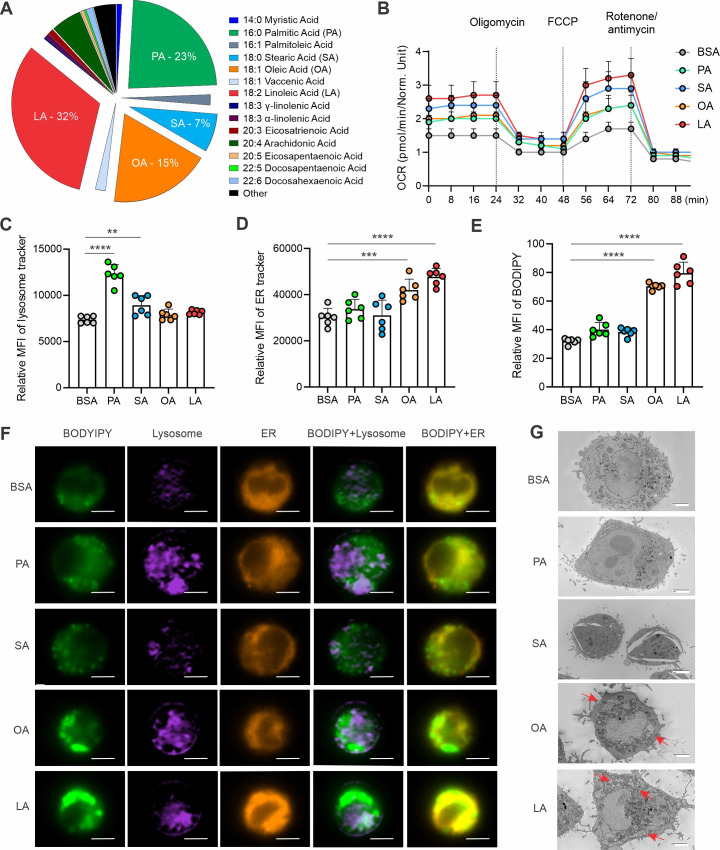
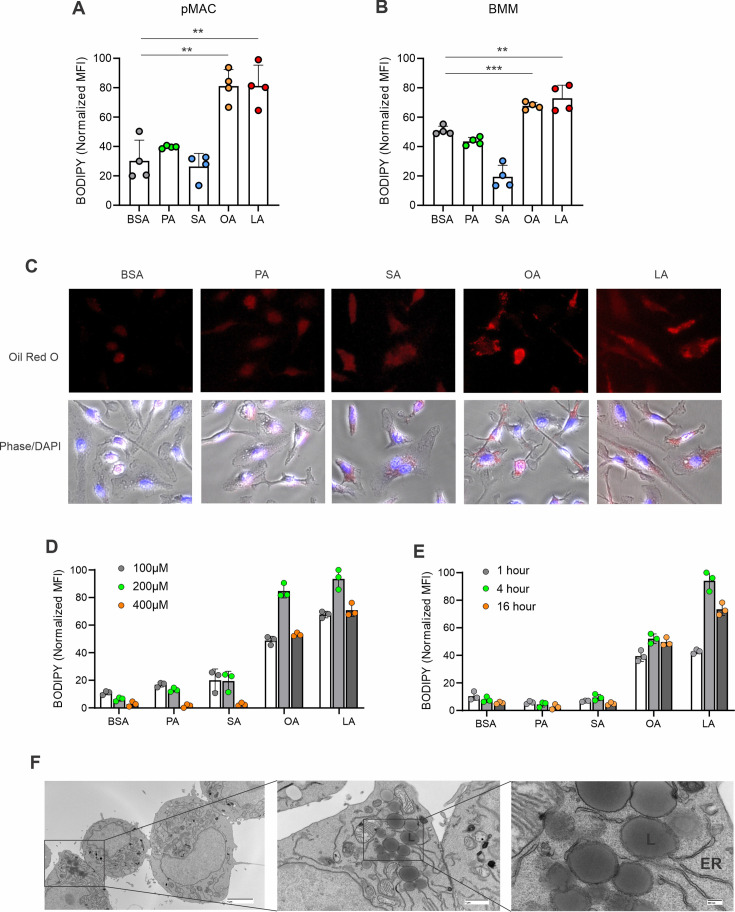
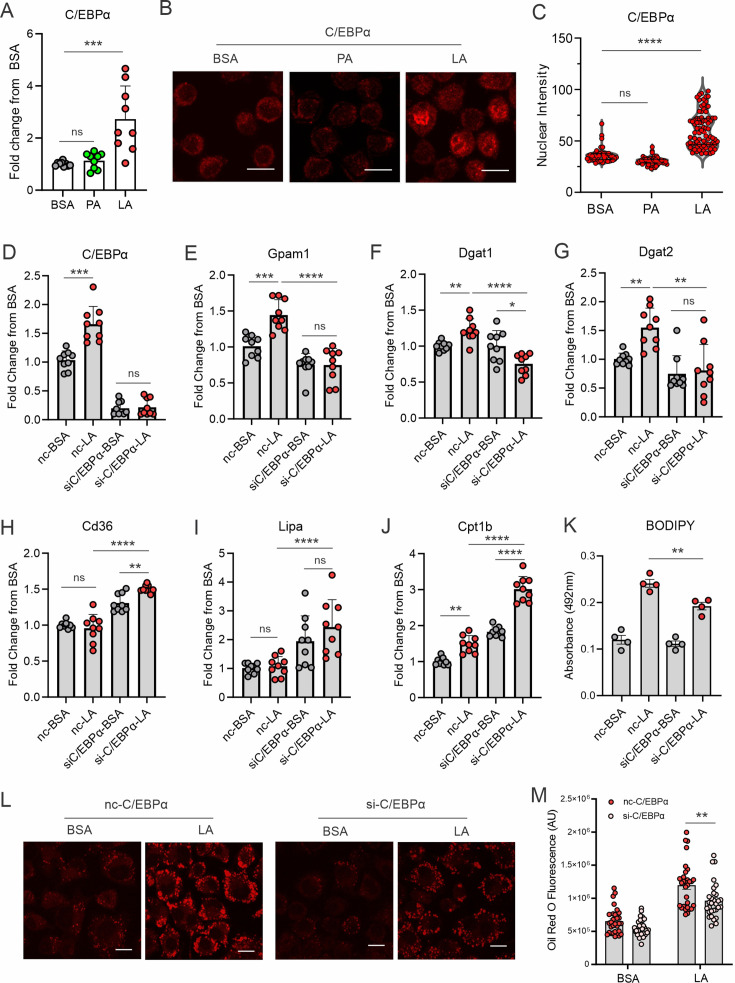
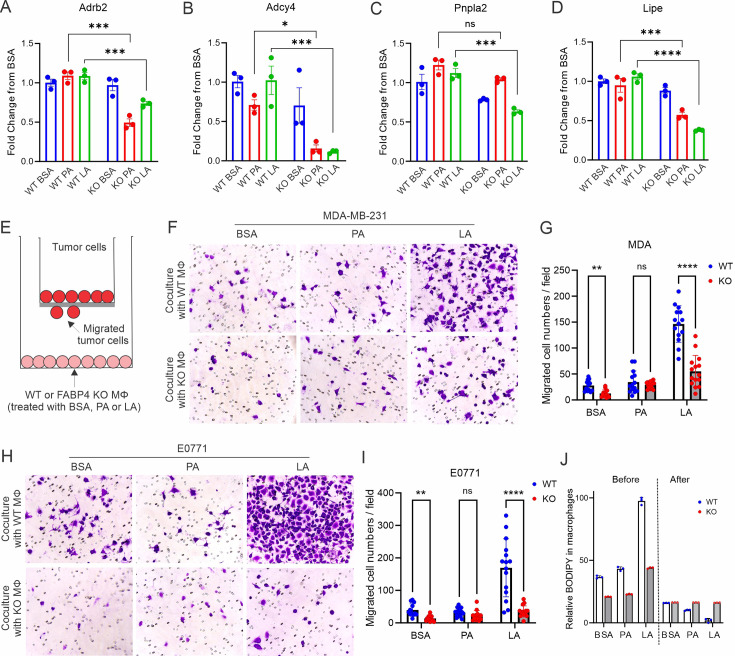
A high density of tumor-associated macrophages (TAMs) is associated with poorer prognosis and survival in breast cancer patients. Recent studies have shown that lipid accumulation in TAMs can promote tumor growth and metastasis in various models. However, the specific molecular mechanisms that drive lipid accumulation and tumor progression in TAMs remain largely unknown. Herein, we demonstrated that unsaturated fatty acids (FAs), unlike saturated ones, are more likely to form lipid droplets in murine macrophages. Specifically, unsaturated FAs, including linoleic acids (LA), activate the FABP4/CEBPα pathway, leading to triglyceride synthesis and lipid droplet formation. Furthermore, FABP4 enhances lipolysis and FA utilization by breast cancer cell lines, which promotes cancer cell migration in vitro and metastasis in vivo. Notably, a deficiency of FABP4 in murine macrophages significantly reduces LA-induced lipid metabolism. Therefore, our findings suggest FABP4 as a crucial lipid messenger that facilitates unsaturated FA-mediated lipid accumulation and lipolysis in TAMs, thus contributing to the metastasis of breast cancer.
Keywords: FABP4; breast cancer; cancer biology; human; lipid droplet formation; metastasis; mouse; tumor associated macrophages; unsaturated fats.
Conflict of interest statement
MY, XJ, SL, JH, JY, AA, ZL, MC, HK, JS, AK, MK, YX, ES, SS, BL No competing interests declared
Figures











Update of
-
FABP4-mediated lipid accumulation and lipolysis in tumor associated macrophages promote breast cancer metastasis.bioRxiv [Preprint]. 2024 Jul 4:2024.07.02.601733. doi: 10.1101/2024.07.02.601733. bioRxiv. 2024. Update in: Elife. 2024 Nov 08;13:RP101221. doi: 10.7554/eLife.101221. PMID: 39005322 Free PMC article. Updated. Preprint.
References
-
- Chen D, Wirth KM, Kizy S, Muretta JM, Markowski TW, Yong P, Sheka A, Abdelwahab H, Hertzel AV, Ikramuddin S, Yamamoto M, Bernlohr DA. Desmoglein 2 functions as a receptor for fatty acid binding protein 4 in breast cancer epithelial cells. Molecular Cancer Research. 2023;21:836–848. doi: 10.1158/1541-7786.MCR-22-0763. - DOI - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
