

SUMOylation of rice DELLA SLR1 modulates transcriptional responses and improves yield under salt stress
- PMID: 39514093
- PMCID: PMC11549141
- DOI: 10.1007/s00425-024-04565-1
SUMOylation of rice DELLA SLR1 modulates transcriptional responses and improves yield under salt stress
Abstract
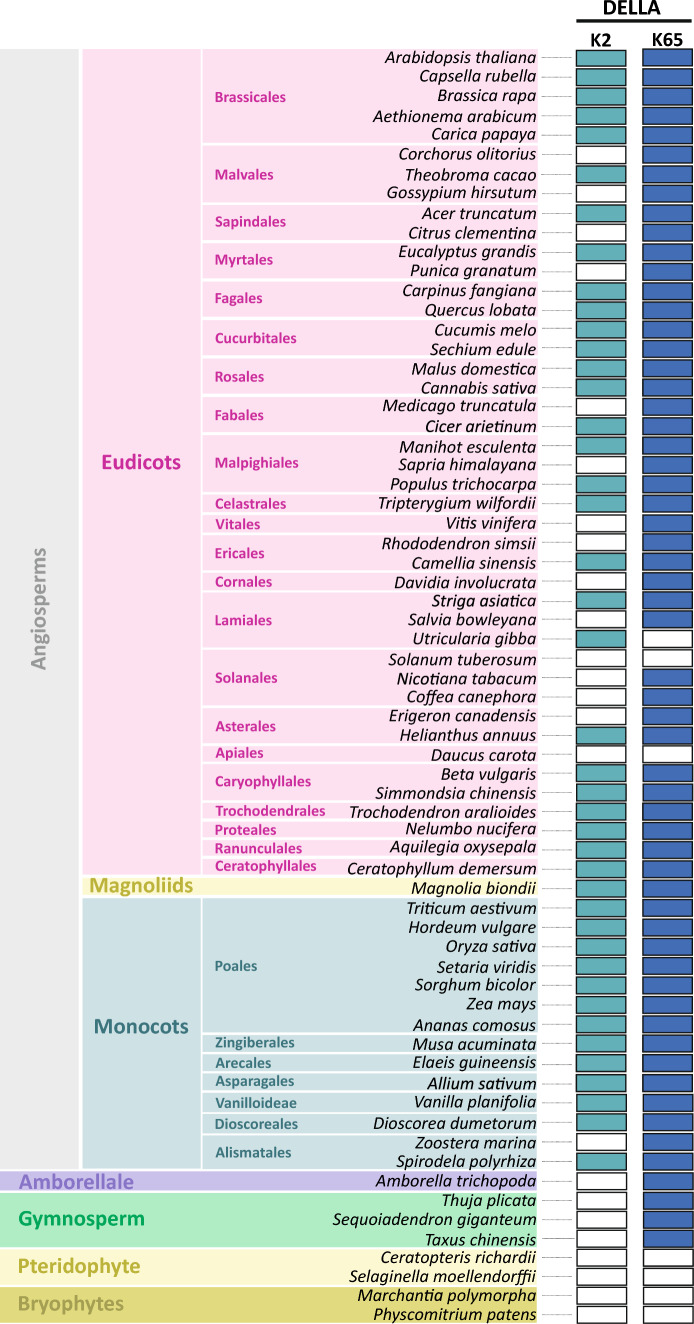
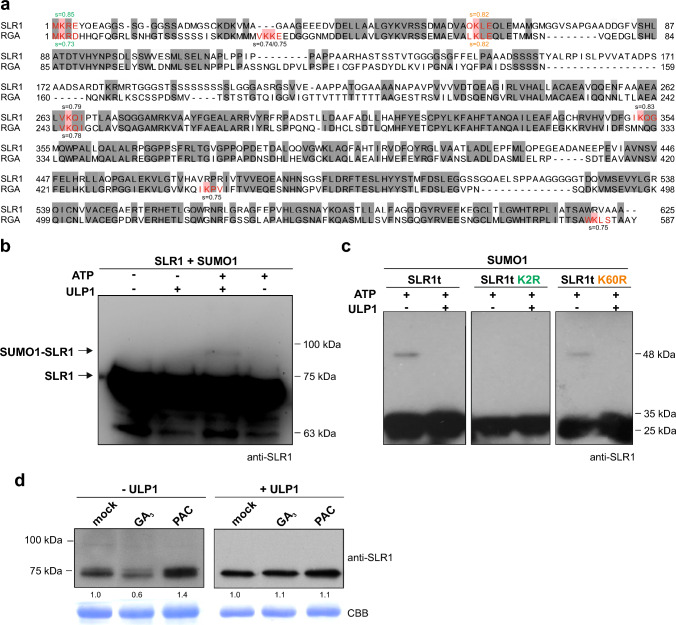
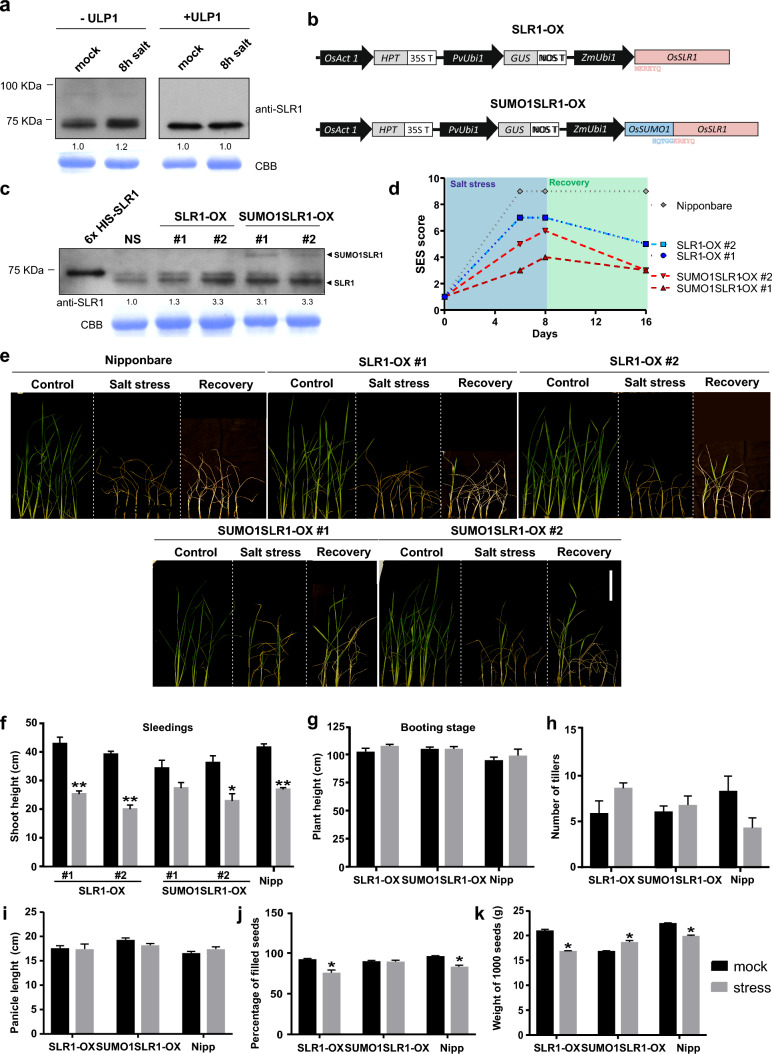
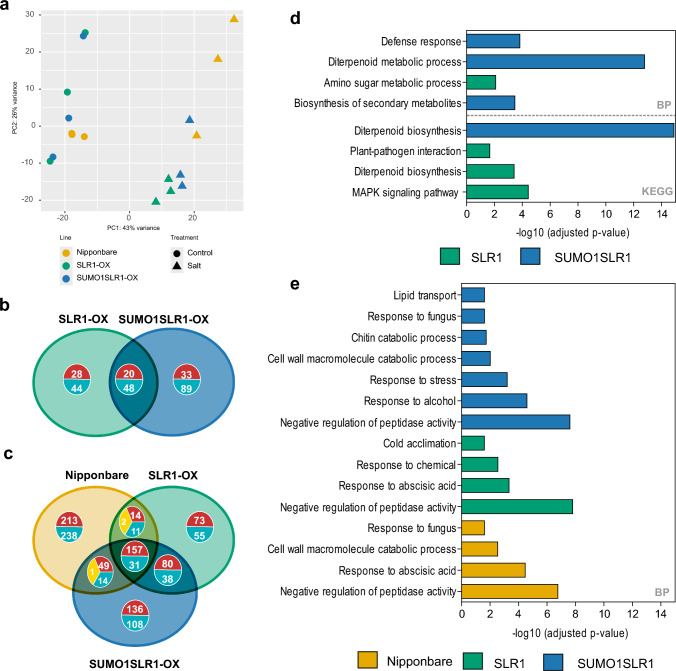
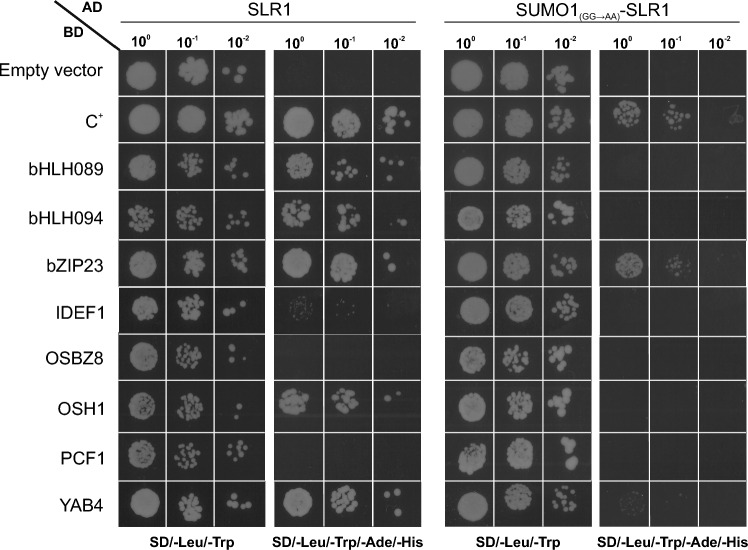
SUMOylation of SLR1 at K2 protects productivity under salt stress, possibly by modulation of SLR1 interactome. DELLA proteins modulate GA signaling and are major regulators of plant plasticity to endure stress. DELLAs are mostly regulated at the post-translational level, and their activity relies on the interaction with upstream regulators and transcription factors (TFs). SUMOylation is a post-translational modification (PTM) capable of changing protein interaction and has been found to influence DELLA activity in Arabidopsis. We determined that SUMOylation of the single rice DELLA, SLENDER RICE1 (SLR1), occurs in a lysine residue different from the one identified in Arabidopsis REPRESSOR OF GA (RGA). Artificially increasing the SUMOylated SLR1 levels attenuated the penalty of salt stress on rice yield. Gene expression analysis revealed that the overexpression of SUMOylated SLR1 can regulate GA biosynthesis, which could partially explain the sustained productivity upon salt stress imposition. Furthermore, SLR1 SUMOylation blocked the interaction with the growth regulator YAB4, which may fine-tune GA20ox2 expression. We also identified novel SLR1 interactors: bZIP23, bHLH089, bHLH094, and OSH1. All those interactions were impaired in the presence of SUMOylated SLR1. Mechanistically, we propose that SUMOylation of SLR1 disrupts its interaction with several transcription factors implicated in GA-dependent growth and ABA-dependent salinity tolerance to modulate downstream gene expression. We found that SLR1 SUMOylation represents a novel mechanism modulating DELLA activity, which attenuates the impact of stress on plant performance.
Keywords: Oryza sativa; DELLA; Gibberellin; Productivity; SLR1; SUMOylation; Salt stress; Transcription factors.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Characterization of the molecular mechanism underlying gibberellin perception complex formation in rice.Plant Cell. 2010 Aug;22(8):2680-96. doi: 10.1105/tpc.110.075549. Epub 2010 Aug 17. Plant Cell. 2010. PMID: 20716699 Free PMC article.
-
The DELLA Protein SLR1 Integrates and Amplifies Salicylic Acid- and Jasmonic Acid-Dependent Innate Immunity in Rice.Plant Physiol. 2016 Mar;170(3):1831-47. doi: 10.1104/pp.15.01515. Epub 2016 Feb 1. Plant Physiol. 2016. PMID: 26829979 Free PMC article.
-
Release of the repressive activity of rice DELLA protein SLR1 by gibberellin does not require SLR1 degradation in the gid2 mutant.Plant Cell. 2008 Sep;20(9):2437-46. doi: 10.1105/tpc.108.061648. Epub 2008 Sep 30. Plant Cell. 2008. PMID: 18827181 Free PMC article.
-
Gibberellin receptor and its role in gibberellin signaling in plants.Annu Rev Plant Biol. 2007;58:183-98. doi: 10.1146/annurev.arplant.58.032806.103830. Annu Rev Plant Biol. 2007. PMID: 17472566 Review.
-
Regulation of DELLA Proteins by Post-translational Modifications.Plant Cell Physiol. 2020 Dec 23;61(11):1891-1901. doi: 10.1093/pcp/pcaa113. Plant Cell Physiol. 2020. PMID: 32886774 Free PMC article. Review.
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311:91–94. 10.1126/science.1118642 - PubMed
-
- Almadanim MC, Alexandre BM, Rosa MTG, Sapeta H, Leitão AE, Ramalho JC, Lam TT, Negrão S, Abreu IA, Oliveira MM (2017) Rice calcium-dependent protein kinase OsCPK17 targets plasma membrane intrinsic protein and sucrose-phosphate synthase and is required for a proper cold stress response. Plant Cell Environ 40:1197–1213. 10.1111/pce.12916 - PubMed
-
- Camut L, Davière JM, Achard P (2017) Dynamic regulation of DELLA protein activity: SPINDLY and SECRET AGENT unmasked! Mol Plant 10:785–787. 10.1016/j.molp.2017.05.003 - PubMed
MeSH terms
Substances
Grants and funding
- SFRH/BD/85799/2012/Fundação para a Ciência e a Tecnologia
- PD/BD/135584/2018/Fundação para a Ciência e a Tecnologia
- DOI: 10.54499/DL57/2016/CP1369/CT0029/Fundação para a Ciência e a Tecnologia
- IF/00764/2014/Fundação para a Ciência e a Tecnologia
- PTDC/BIA-FBT/31211/2017/Fundação para a Ciência e a Tecnologia
- KBBE/0001/2012/Fundação para a Ciência e a Tecnologia
- Fundação para a Ciência e a Tecnologia/Fundação para a Ciência e a Tecnologia
- UIDP/Multi/04551/2013/Fundação para a Ciência e a Tecnologia
- DOI 10.54499/UIDB/04551/2020/Fundação para a Ciência e a Tecnologia
- DOI 10.54499/UIDP/04551/2020/Fundação para a Ciência e a Tecnologia
LinkOut - more resources
Full Text Sources
Research Materials
