

Mechanisms of rapid plant community change from the Miocene Succor Creek flora, Oregon and Idaho (USA)
- PMID: 39514583
- PMCID: PMC11548735
- DOI: 10.1371/journal.pone.0312104
Mechanisms of rapid plant community change from the Miocene Succor Creek flora, Oregon and Idaho (USA)
Abstract
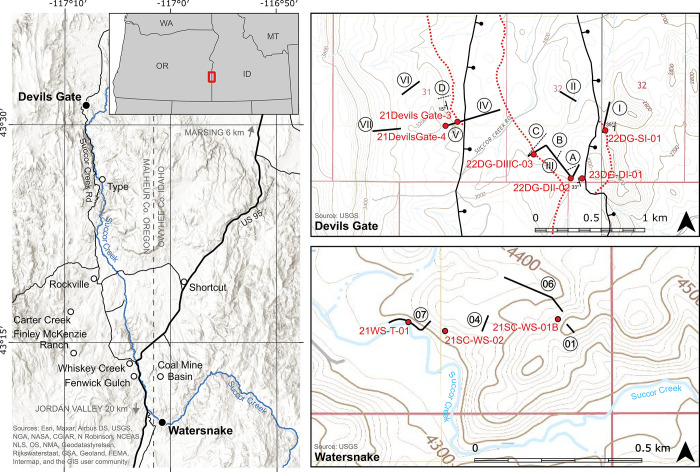
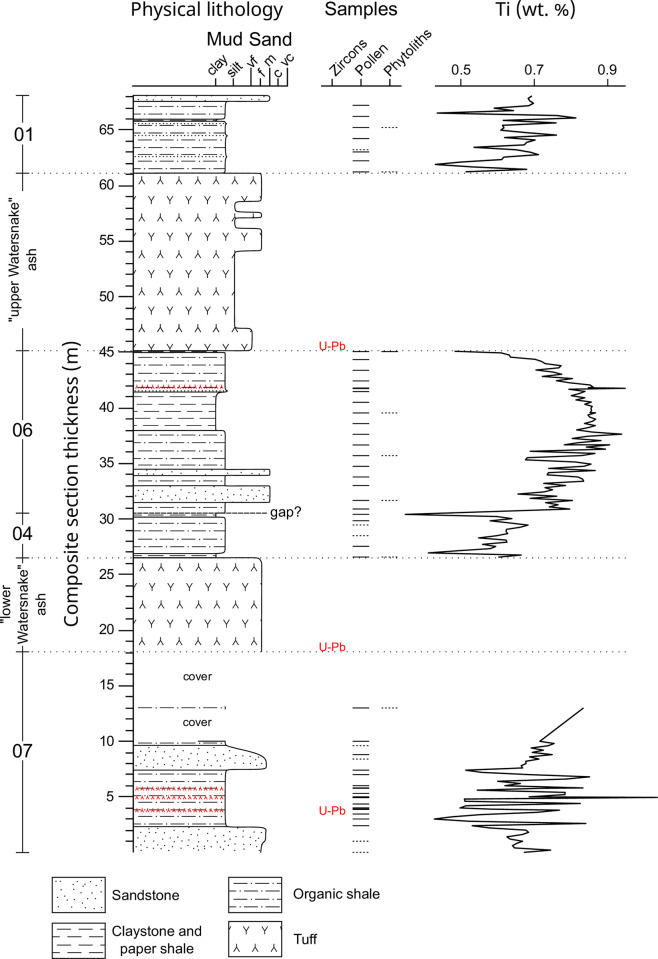
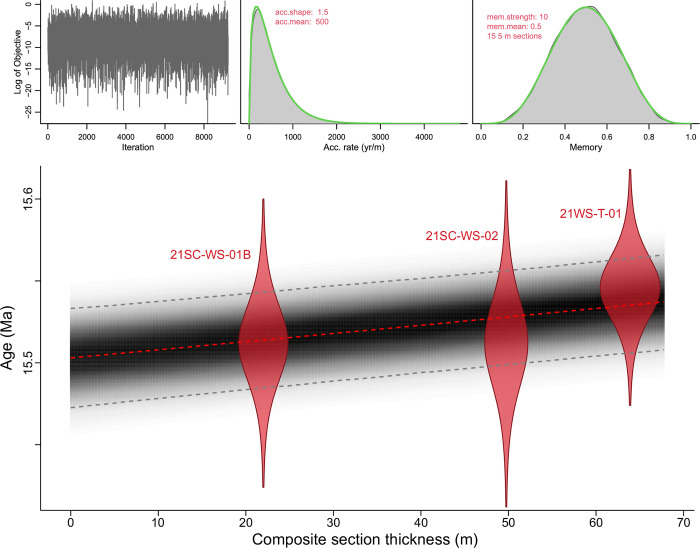
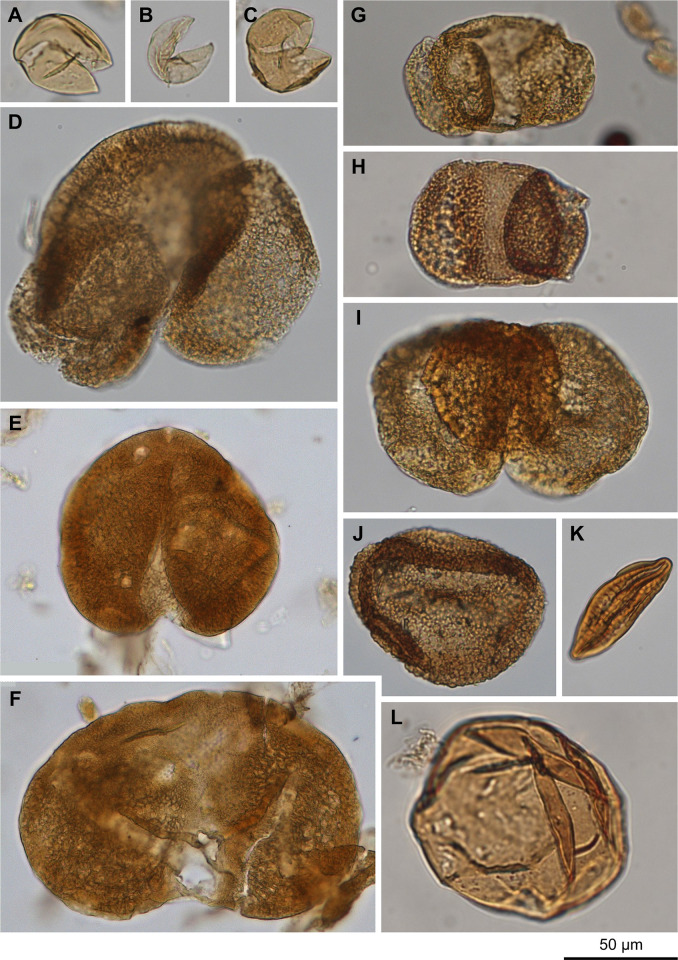
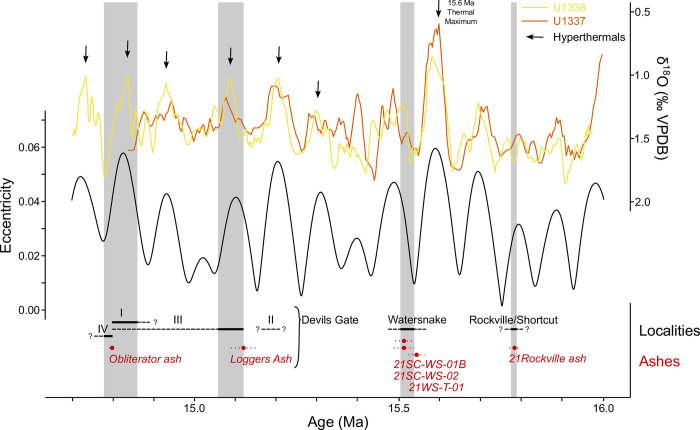
The fossil record of the U.S. Pacific Northwest preserves many Middle Miocene floras with potential for revealing long-term climate-vegetation dynamics during the Miocene Climatic Optimum. However, the possibility of strong, eccentricity-paced climate oscillations and concurrent, intense volcanism may obscure the signature of prevailing, long-term Miocene climate change. To test the hypothesis that volcanic disturbance drove Middle Miocene vegetation dynamics, high-resolution, stratigraphic pollen records and other paleobotanical data from nine localities of the Sucker Creek Formation were combined with sedimentological and geochemical evidence of disturbance within an updated chronostratigraphic framework based on new U-Pb zircon ages from tuffs. The new ages establish a refined, minimum temporal extent of the Sucker Creek Formation, ~15.8 to ~14.8 Ma, and greatly revise the local and regional chronostratigraphic correlations of its dispersed outcrop belt. Our paleoecological analysis at one ~15.52 Ma locality reveals two abrupt shifts in pollen spectra coinciding with the deposition of thick ash-flow tuffs, wherein vegetation dominated by Cupressaceae/Taxaceae, probably representing a Glyptostrobus oregonensis swamp, and upland conifers was supplanted by early-successional forests with abundant Alnus and Betula. Another ephemeral shift from Cupressaceae/Taxaceae swamp taxa in favor of upland conifers Pinus and Tsuga correlates with a shift from low-Ti shale to high-Ti claystone, suggesting a link between altered surface hydrology and vegetation. In total, three rapid vegetation shifts coincide with ash-flow tuffs and are attributed to volcanic disturbance. Longer-term variability between localities, spanning ~1 Myr of the Miocene Climatic Optimum, is chiefly attributed to eccentricity-paced climate change. Overall, Succor Creek plant associations changed frequently over ≤105 years timespans, reminiscent of Quaternary vegetation records. Succor Creek stratigraphic palynology suggests that numerous and extensive collection of stratigraphically controlled samples is necessary to understand broader vegetation trends through time.
Copyright: © 2024 Schiller et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

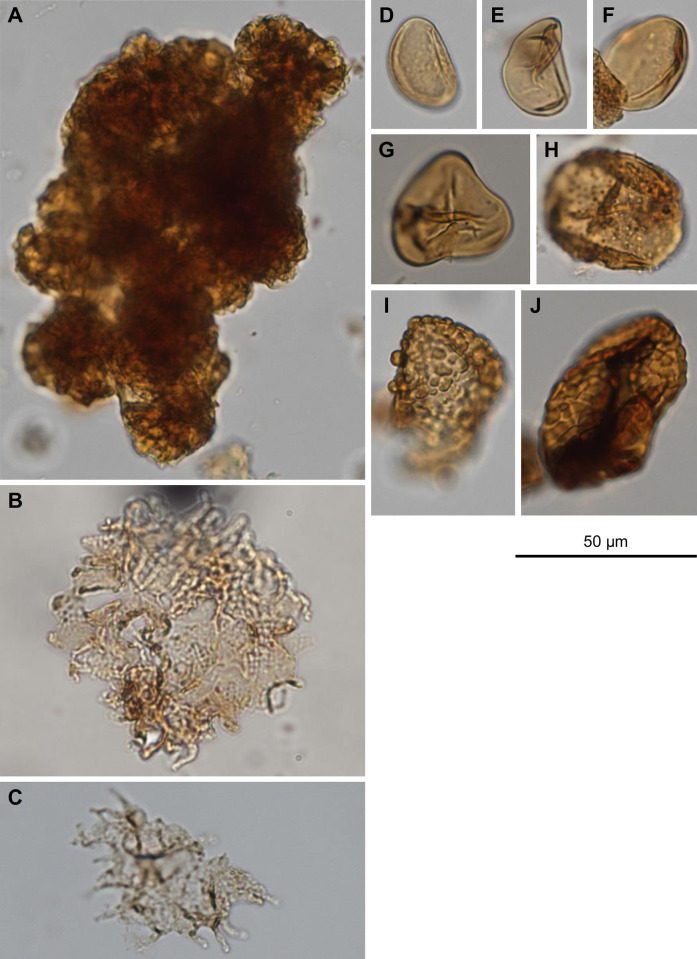
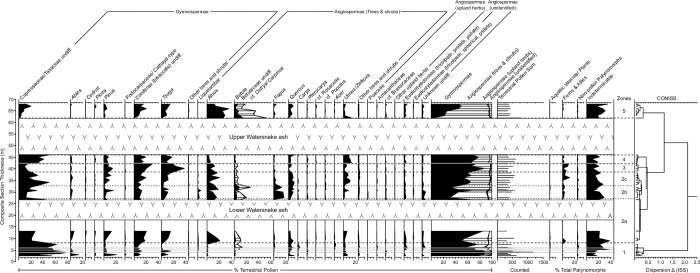
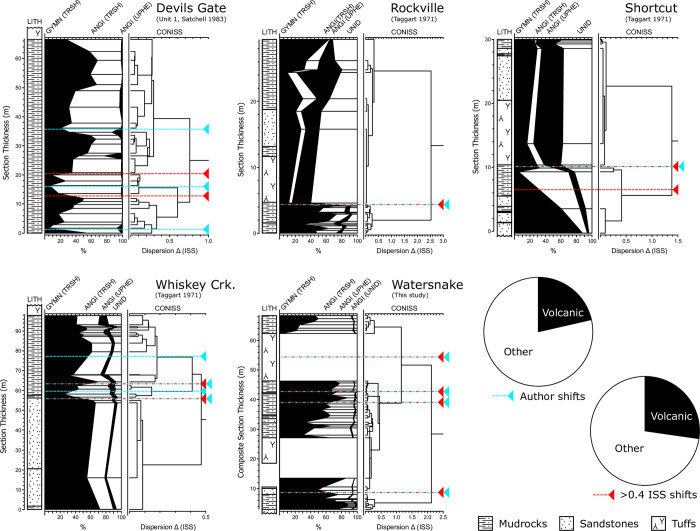
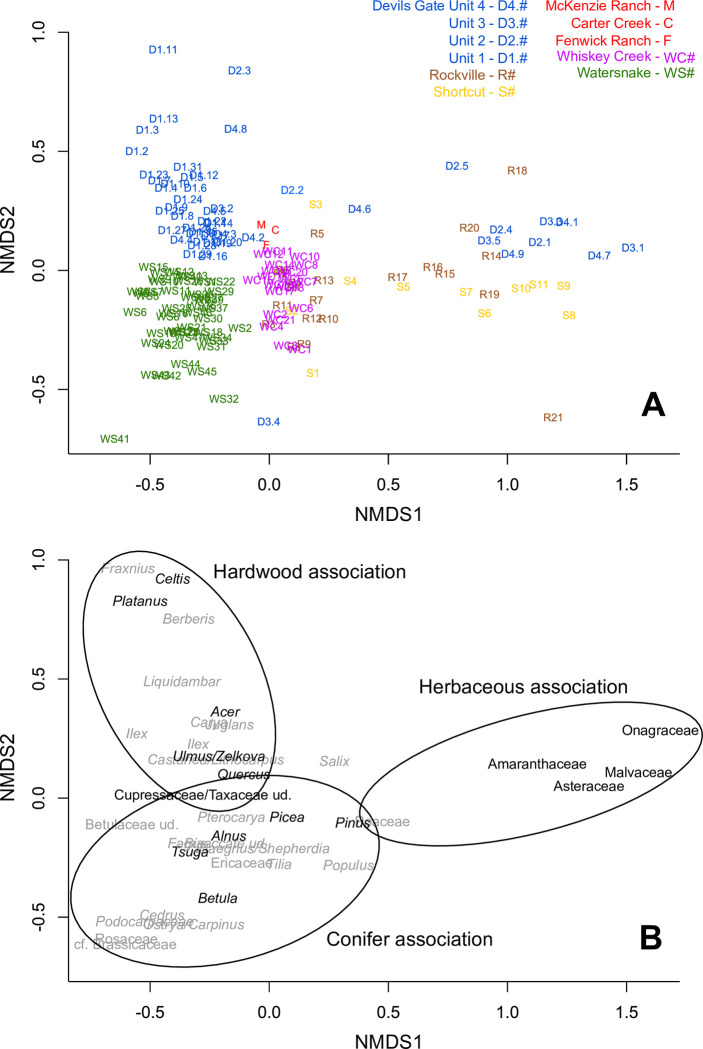
References
-
- Steinthorsdottir M, Coxall HK, de Boer AM, Huber M, Barbolini N, Bradshaw CD, et al.. The Miocene: The Future of the Past. Paleoceanogr Paleoclimatology. 2021;36(4):e2020PA004037.
-
- Goldner A, Herold N, Huber M. The challenge of simulating the warmth of the mid-Miocene climatic optimum in CESM1. Clim Past. 2014. Mar 13;10(2):523–36.
-
- Burls NJ, Bradshaw CD, De Boer AM, Herold N, Huber M, Pound M, et al.. Simulating Miocene Warmth: Insights From an Opportunistic Multi-Model Ensemble (MioMIP1). Paleoceanogr Paleoclimatology. 2021;36(5):e2020PA004054.
-
- Wolfe JA. Distribution of Major Vegetational Types During the Tertiary. In: Sundquist ET, Broecker WS, editors. The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present [Internet]. Washington, D. C.: American Geophysical Union; 1985. [cited 2023 Feb 15]. p. 357–75. (Geophysical Monograph Series; vol. 32). Available from: http://doi.wiley.com/10.1029/GM032p0357 - DOI
-
- Pound MJ, Haywood AM, Salzmann U, Riding JB. Global vegetation dynamics and latitudinal temperature gradients during the Mid to Late Miocene (15.97–5.33Ma). Earth-Sci Rev. 2012. Apr;112(1–2):1–22.
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
