

VTA dopamine neurons are hyperexcitable in 3xTg-AD mice due to casein kinase 2-dependent SK channel dysfunction
- PMID: 39516200
- PMCID: PMC11549218
- DOI: 10.1038/s41467-024-53891-1
VTA dopamine neurons are hyperexcitable in 3xTg-AD mice due to casein kinase 2-dependent SK channel dysfunction
Abstract
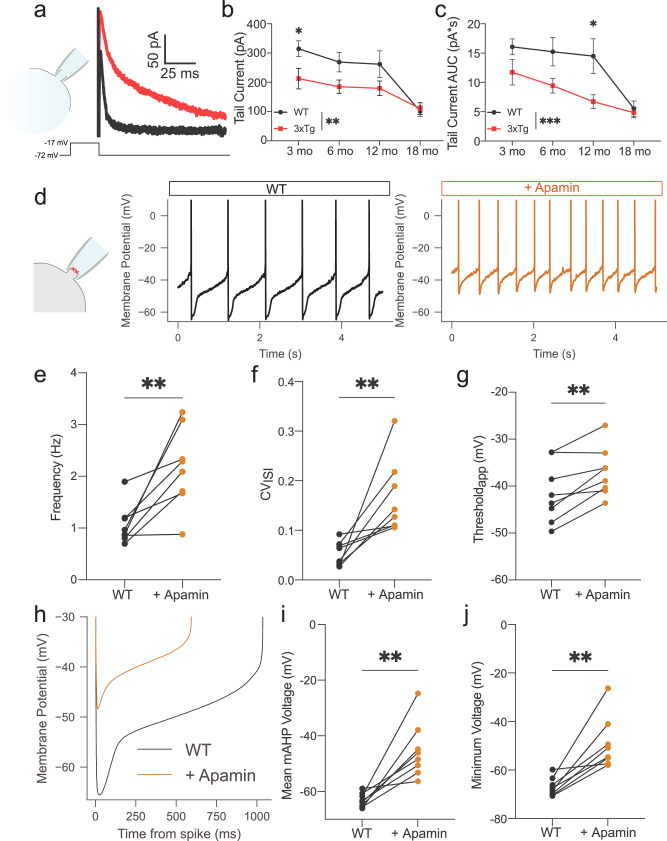
Alzheimer's disease (AD) patients exhibit neuropsychiatric symptoms that extend beyond classical cognitive deficits, suggesting involvement of subcortical areas. Here, we investigated the role of midbrain dopamine (DA) neurons in AD using the amyloid + tau-driven 3xTg-AD mouse model. We found deficits in reward-based operant learning in AD mice, suggesting possible VTA DA neuron dysregulation. Physiological assessment revealed hyperexcitability and disrupted firing in DA neurons caused by reduced activity of small-conductance calcium-activated potassium (SK) channels. RNA sequencing from contents of single patch-clamped DA neurons (Patch-seq) identified up-regulation of the SK channel modulator casein kinase 2 (CK2), which we corroborated by immunohistochemical protein analysis. Pharmacological inhibition of CK2 restored SK channel activity and normal firing patterns in 3xTg-AD mice. These findings identify a mechanism of ion channel dysregulation in VTA DA neurons that could contribute to behavioral abnormalities in AD, paving the way for novel treatment strategies.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures







Update of
-
VTA dopamine neurons are hyperexcitable in 3xTg-AD mice due to casein kinase 2-dependent SK channel dysfunction.bioRxiv [Preprint]. 2023 Nov 17:2023.11.16.567486. doi: 10.1101/2023.11.16.567486. bioRxiv. 2023. Update in: Nat Commun. 2024 Nov 8;15(1):9673. doi: 10.1038/s41467-024-53891-1. PMID: 38014232 Free PMC article. Updated. Preprint.
References
-
- La Barbera, L. et al. Nilotinib restores memory function by preventing dopaminergic neuron degeneration in a mouse model of Alzheimer’s Disease. Prog. Neurobiol.202, 102031 (2021). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- P30 AG050911, R01 AG059430/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- I01 BX005396/BX/BLRD VA/United States
- R21 AG072811, R01 AG052606/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R21 AG072811/AG/NIA NIH HHS/United States
- F31 AG079620/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
