

Structural basis of 3'-tRNA maturation by the human mitochondrial RNase Z complex
- PMID: 39516281
- PMCID: PMC11649783
- DOI: 10.1038/s44318-024-00297-w
Structural basis of 3'-tRNA maturation by the human mitochondrial RNase Z complex
Abstract
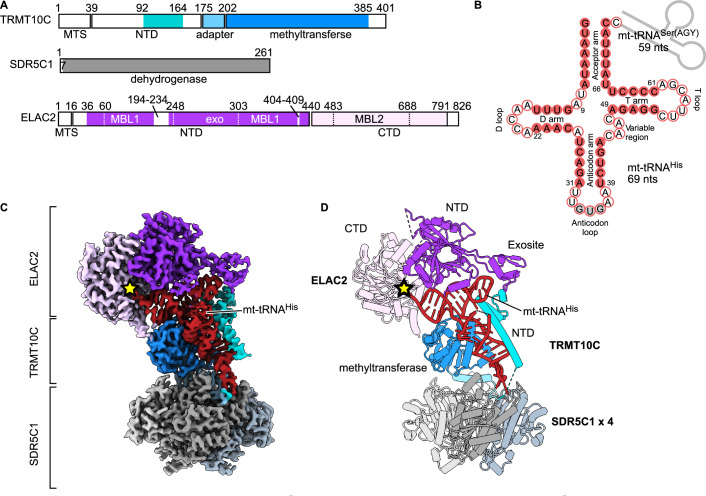
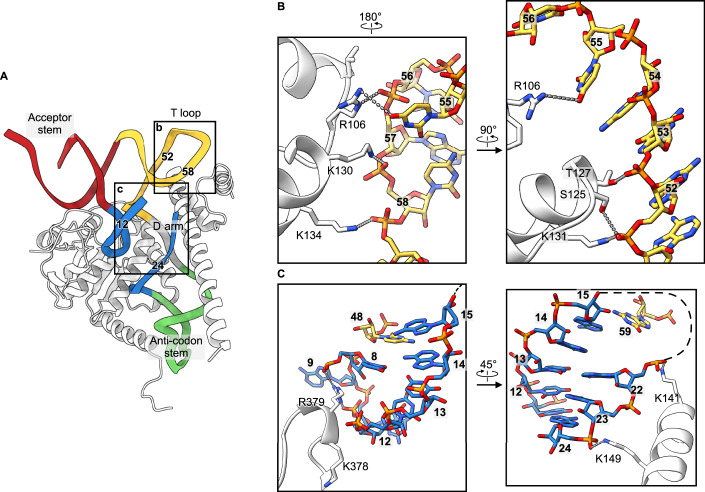
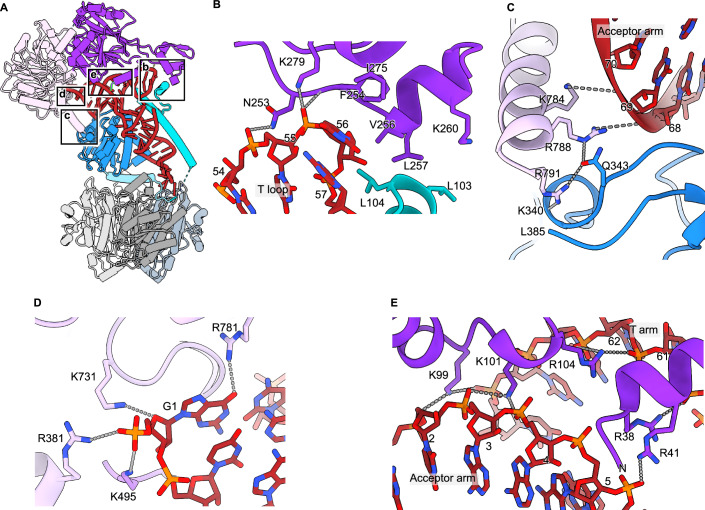
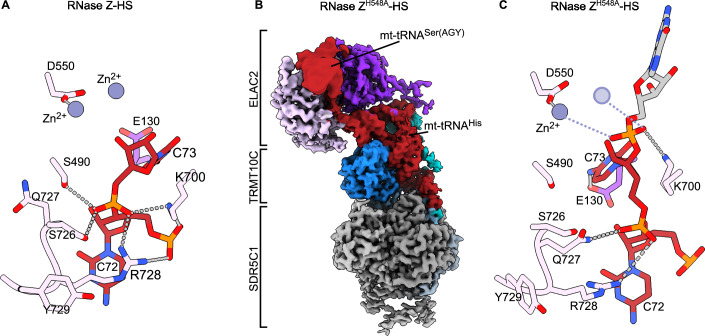
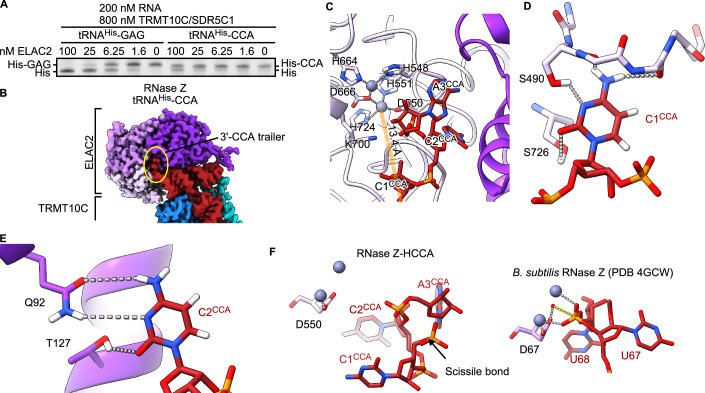
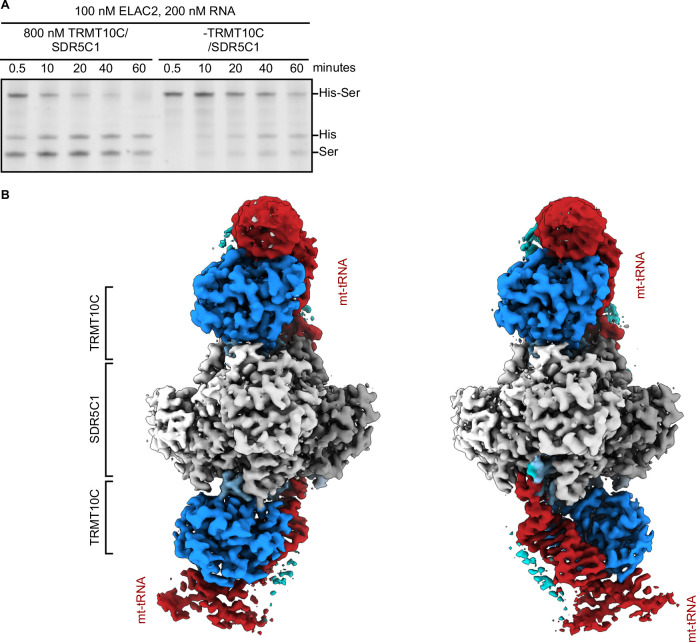
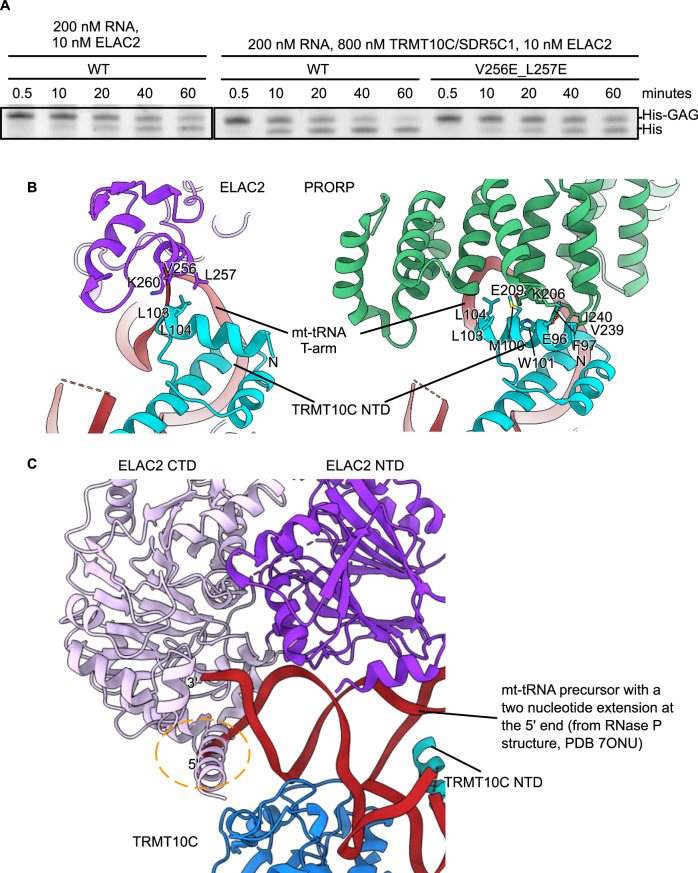
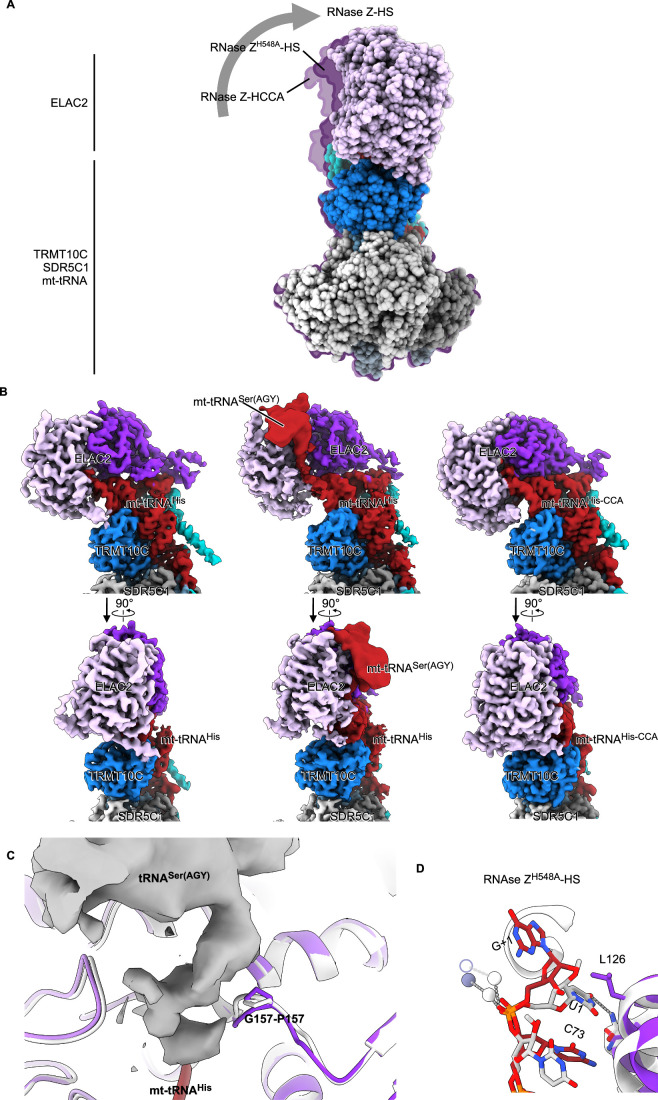
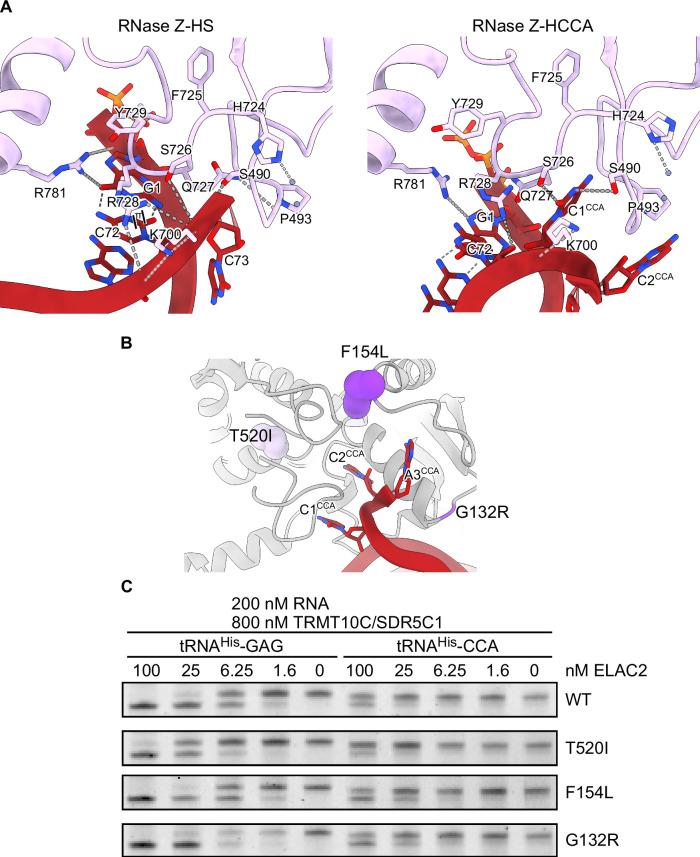
Maturation of human mitochondrial tRNA is essential for cellular energy production, yet the underlying mechanisms remain only partially understood. Here, we present several cryo-EM structures of the mitochondrial RNase Z complex (ELAC2/SDR5C1/TRMT10C) bound to different maturation states of mitochondrial tRNAHis, showing the molecular basis for tRNA-substrate selection and catalysis. Our structural insights provide a molecular rationale for the 5'-to-3' tRNA processing order in mitochondria, the 3'-CCA antideterminant effect, and the basis for sequence-independent recognition of mitochondrial tRNA substrates. Furthermore, our study links mutations in ELAC2 to clinically relevant mitochondrial diseases, offering a deeper understanding of the molecular defects contributing to these conditions.
Keywords: Cryo-EM; ELAC2; Mitochondria; RNA Processing; RNase Z.
© 2024. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The authors declare no competing interests.
Figures
Similar articles
-
Molecular basis of human nuclear and mitochondrial tRNA 3' processing.Nat Struct Mol Biol. 2025 Apr;32(4):613-624. doi: 10.1038/s41594-024-01445-w. Epub 2025 Jan 2. Nat Struct Mol Biol. 2025. PMID: 39747487 Free PMC article.
-
Advances in Human Pre-tRNA Maturation: TRMT10C and ELAC2 in Focus.J Mol Biol. 2025 Aug 15;437(16):168989. doi: 10.1016/j.jmb.2025.168989. Epub 2025 Feb 10. J Mol Biol. 2025. PMID: 39938738 Review.
-
Structural basis for human mitochondrial tRNA maturation.Nat Commun. 2024 Jun 1;15(1):4683. doi: 10.1038/s41467-024-49132-0. Nat Commun. 2024. PMID: 38824131 Free PMC article.
-
Disease-associated mutations in mitochondrial precursor tRNAs affect binding, m1R9 methylation, and tRNA processing by mtRNase P.RNA. 2021 Apr;27(4):420-432. doi: 10.1261/rna.077198.120. Epub 2020 Dec 30. RNA. 2021. PMID: 33380464 Free PMC article.
-
Mitochondrial tRNA modifications: functions, diseases caused by their loss, and treatment strategies.RNA. 2025 Feb 19;31(3):382-394. doi: 10.1261/rna.080257.124. RNA. 2025. PMID: 39719325 Free PMC article. Review.
Cited by
-
FASTKD5 processes mitochondrial pre-mRNAs at noncanonical cleavage sites.Nucleic Acids Res. 2025 Jul 8;53(13):gkaf665. doi: 10.1093/nar/gkaf665. Nucleic Acids Res. 2025. PMID: 40637235 Free PMC article.
-
Bi-allelic mutations in FASTKD5 are associated with cytochrome c oxidase deficiency and early- to late-onset Leigh syndrome.Am J Hum Genet. 2025 Jul 3;112(7):1699-1710. doi: 10.1016/j.ajhg.2025.05.007. Epub 2025 Jun 10. Am J Hum Genet. 2025. PMID: 40499538
-
The Vsr-like protein FASTKD4 regulates the stability and polyadenylation of the MT-ND3 mRNA.Nucleic Acids Res. 2025 Feb 8;53(4):gkae1261. doi: 10.1093/nar/gkae1261. Nucleic Acids Res. 2025. PMID: 39727163 Free PMC article.
-
Mitochondrial tRNA processing: a neutral evolutionary ratchet innovation.Trends Biochem Sci. 2025 Jun 27:S0968-0004(25)00126-4. doi: 10.1016/j.tibs.2025.05.008. Online ahead of print. Trends Biochem Sci. 2025. PMID: 40581570
-
Molecular basis of human nuclear and mitochondrial tRNA 3' processing.Nat Struct Mol Biol. 2025 Apr;32(4):613-624. doi: 10.1038/s41594-024-01445-w. Epub 2025 Jan 2. Nat Struct Mol Biol. 2025. PMID: 39747487 Free PMC article.
References
-
- Altman S, Kirsebom L, Talbot S (1993) Recent Studies of Ribonuclease-P. FASEB J 7:7–14 - PubMed
-
- Bartkiewicz M, Gold H, Altman S (1989) Identification and characterization of an RNA molecule that copurifies with RNase P activity from HeLa cells. Genes Dev 3:488–499 - PubMed
-
- Bhatta A, Kuhle B, Yu RD, Spanaus L, Ditter K, Bohnsack KE, Hillen HS (2024) Molecular basis of human nuclear and mitochondrial tRNA 3’-processing. Preprint at 10.1101/2024.04.04.588063
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
