

Tau processing and tau-mediated inflammation differ in human APOEε2 and APOEε4 astrocytes
- PMID: 39524360
- PMCID: PMC11549983
- DOI: 10.1016/j.isci.2024.111163
Tau processing and tau-mediated inflammation differ in human APOEε2 and APOEε4 astrocytes
Abstract
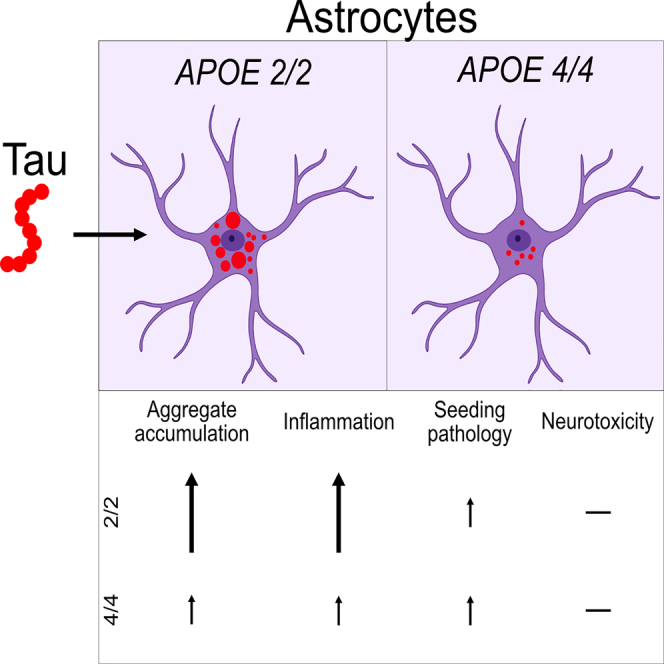
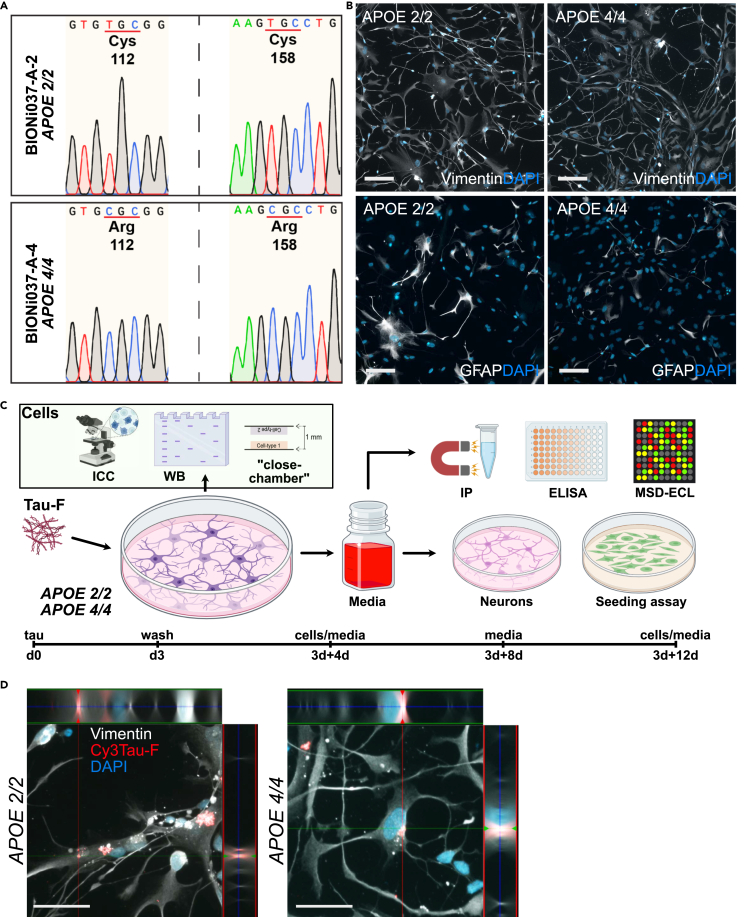
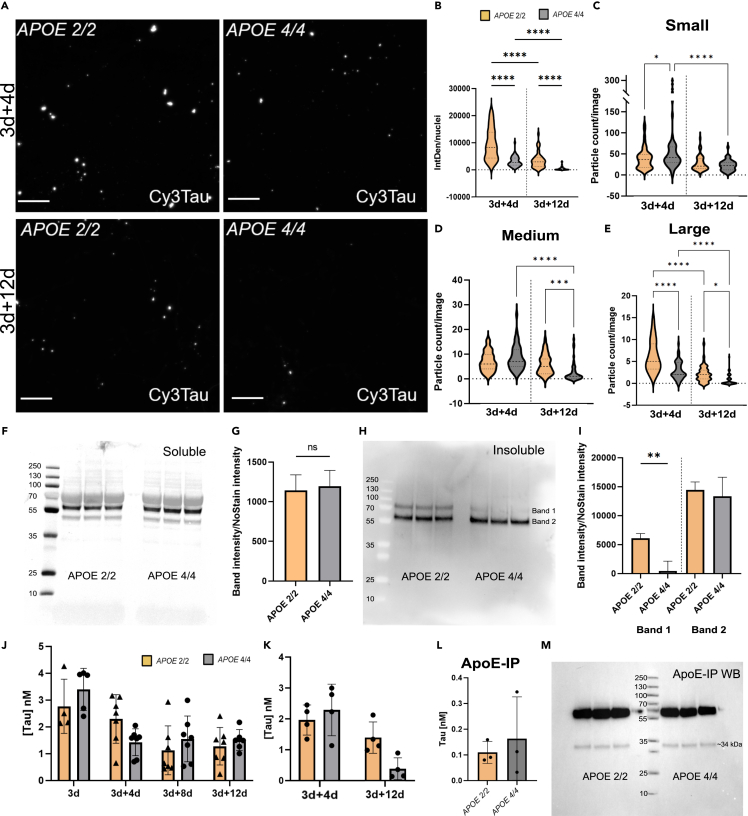
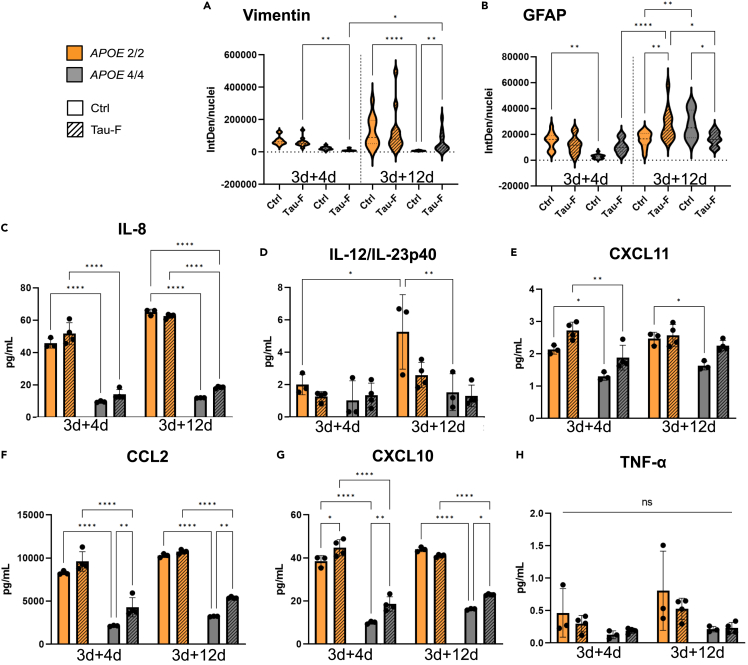
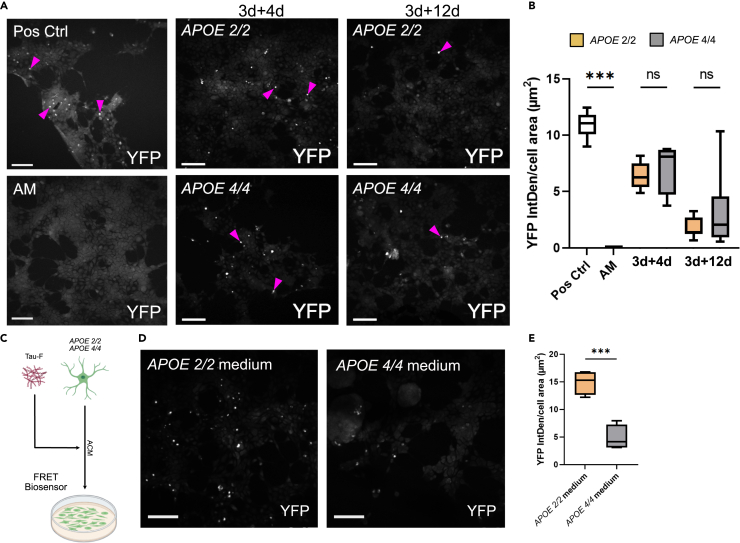
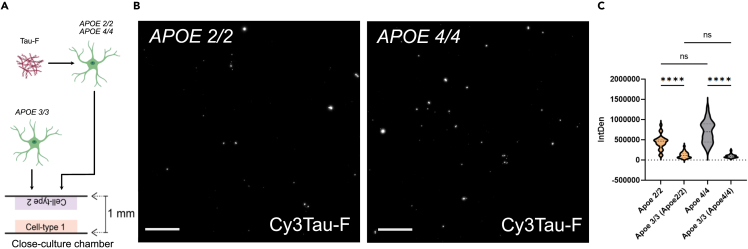
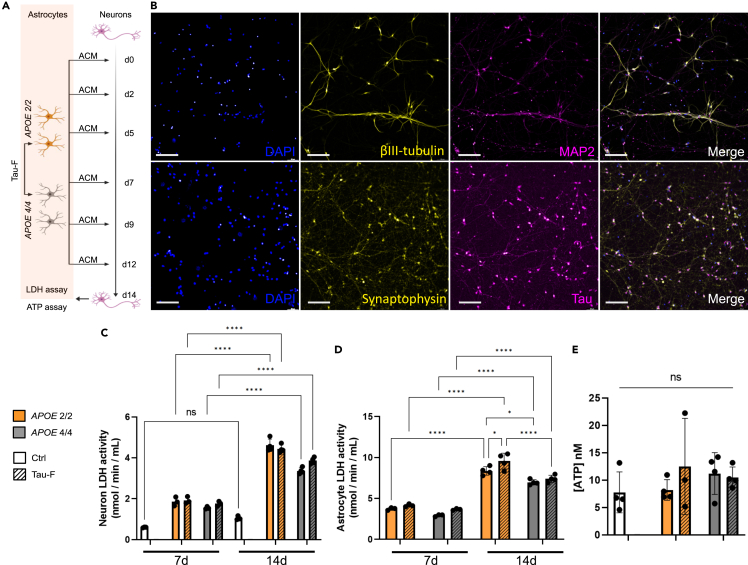
Alzheimer's disease (AD) and progressive supra-nuclear palsy (PSP) are both proteinopathies, characterized by the accumulation of tau aggregates. APOEε4 is the greatest genetic risk factor for developing AD, while APOEε2 is a significant risk factor for developing PSP. In the brain, astrocytes are the predominant producer of ApoE, but they are also important for inflammation and overall brain homeostasis. Although, tau inclusions appear frequently in astrocytes in both AD and PSP brains, their connection to ApoE remains unclear. Here, we show that hiPSC-derived APOE 2/2 astrocytes accumulate, process, and spread pathogenic tau aggregates more efficiently than isogenic APOE 4/4 astrocytes. Moreover, the APOE 2/2 astrocytes display a more robust inflammatory response, which could be of relevance for the disease course. Taken together, our data highlight a central role of ApoE in astrocyte-mediated tau pathology.
Keywords: disease; genes; neuroscience.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that no competing interests.
Figures
Similar articles
-
Astrocytic expression of the Alzheimer's disease risk allele, ApoEε4, potentiates neuronal tau pathology in multiple preclinical models.Sci Rep. 2021 Feb 9;11(1):3438. doi: 10.1038/s41598-021-82901-1. Sci Rep. 2021. PMID: 33564035 Free PMC article.
-
APOE2 orchestrated differences in transcriptomic and lipidomic profiles of postmortem AD brain.Alzheimers Res Ther. 2019 Dec 30;11(1):113. doi: 10.1186/s13195-019-0558-0. Alzheimers Res Ther. 2019. PMID: 31888770 Free PMC article.
-
Pathological tau signatures and nuclear alterations in neurons, astrocytes and microglia in Alzheimer's disease, progressive supranuclear palsy, and dementia with Lewy bodies.Brain Pathol. 2023 Jan;33(1):e13112. doi: 10.1111/bpa.13112. Epub 2022 Aug 24. Brain Pathol. 2023. PMID: 36054524 Free PMC article.
-
Astrocytic inclusions in progressive supranuclear palsy and corticobasal degeneration.Neuropathology. 2014 Dec;34(6):555-70. doi: 10.1111/neup.12143. Epub 2014 Aug 14. Neuropathology. 2014. PMID: 25124031 Review.
-
APOEε4 and risk of Alzheimer's disease - time to move forward.Front Neurosci. 2023 May 19;17:1195724. doi: 10.3389/fnins.2023.1195724. eCollection 2023. Front Neurosci. 2023. PMID: 37274212 Free PMC article. Review.
Cited by
-
The Genetic Background of the Immunological and Inflammatory Aspects of Progressive Supranuclear Palsy.Int J Mol Sci. 2025 Apr 22;26(9):3927. doi: 10.3390/ijms26093927. Int J Mol Sci. 2025. PMID: 40362170 Free PMC article. Review.
References
-
- Komori T., Arai N., Oda M., Nakayama H., Mori H., Yagishita S., Takahashi T., Amano N., Murayama S., Murakami S., et al. Astrocytic plaques and tufts of abnormal fibers do not coexist in corticobasal degeneration and progressive supranuclear palsy. Acta Neuropathol. 1998;96:401–408. doi: 10.1007/s004010050911. - DOI - PubMed
-
- Richetin K., Steullet P., Pachoud M., Perbet R., Parietti E., Maheswaran M., Eddarkaoui S., Bégard S., Pythoud C., Rey M., et al. Tau accumulation in astrocytes of the dentate gyrus induces neuronal dysfunction and memory deficits in Alzheimer’s disease. Nat. Neurosci. 2020;23:1567–1579. doi: 10.1038/s41593-020-00728-x. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
