

N-terminal cleavage of cyclophilin D boosts its ability to bind F-ATP synthase
- PMID: 39528709
- PMCID: PMC11555324
- DOI: 10.1038/s42003-024-07172-8
N-terminal cleavage of cyclophilin D boosts its ability to bind F-ATP synthase
Abstract
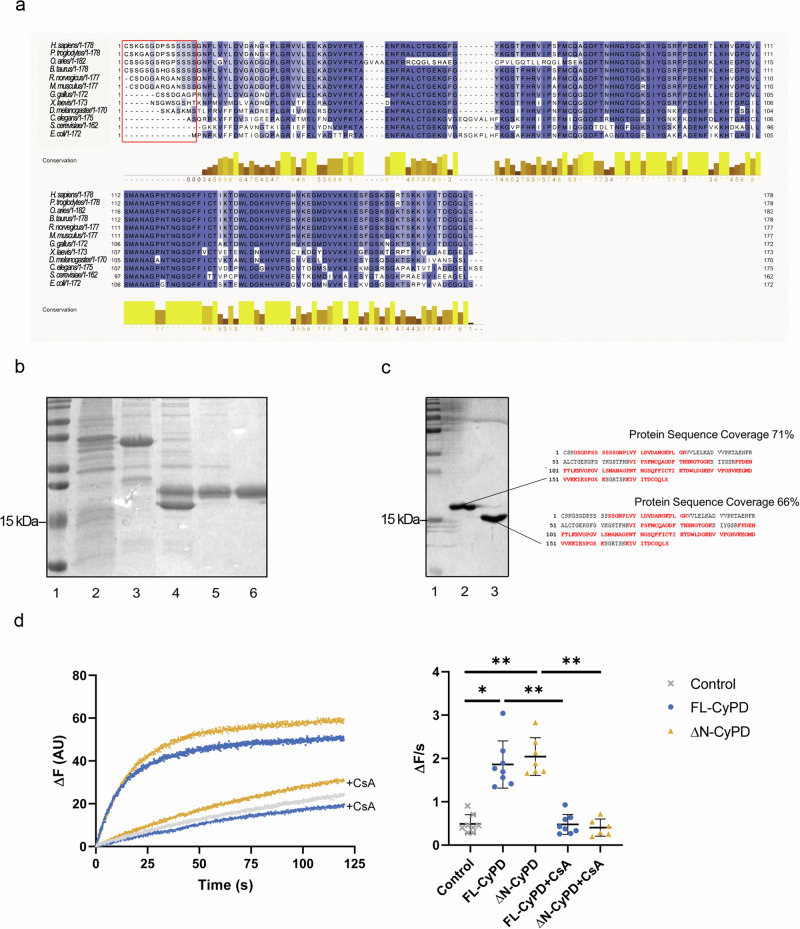
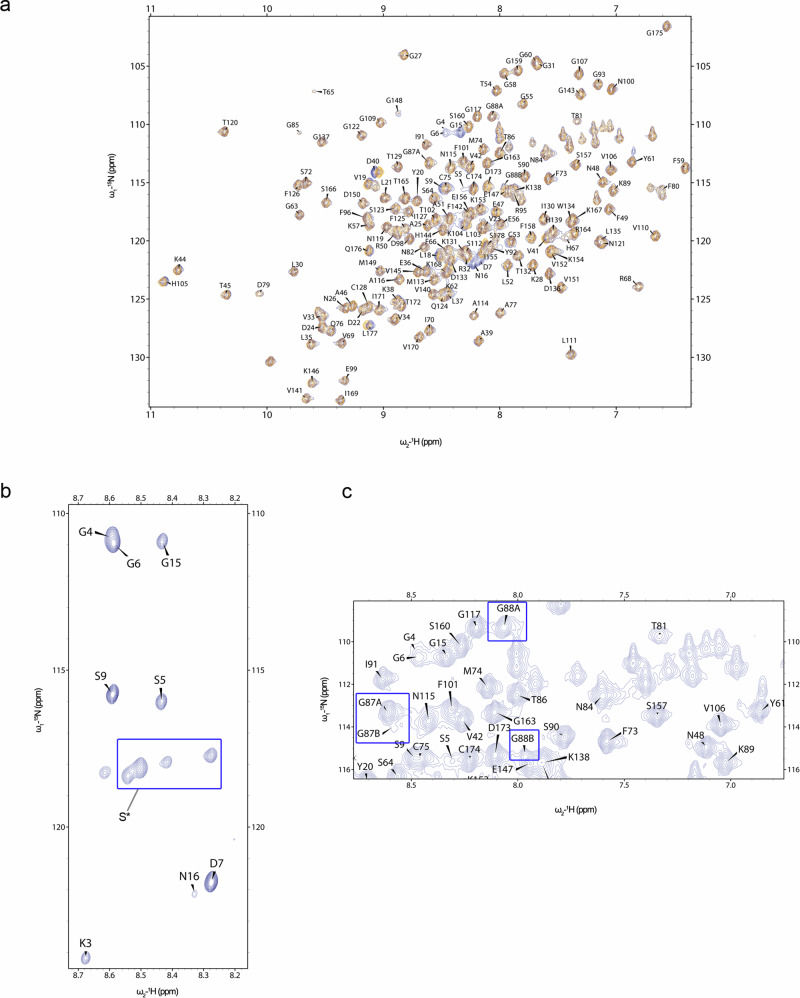
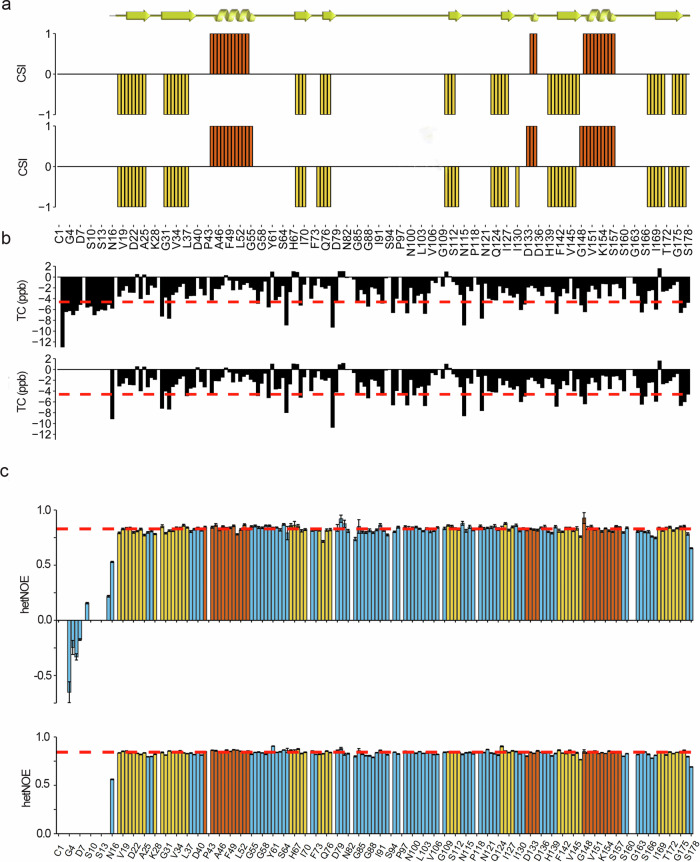
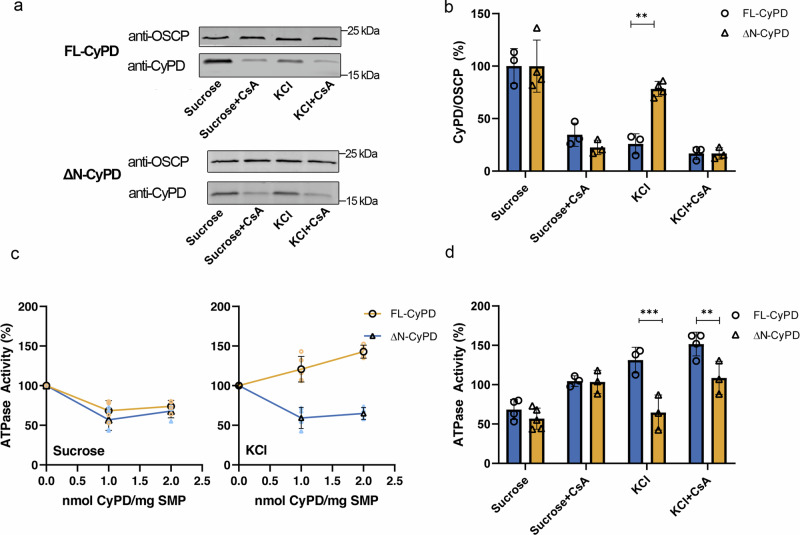
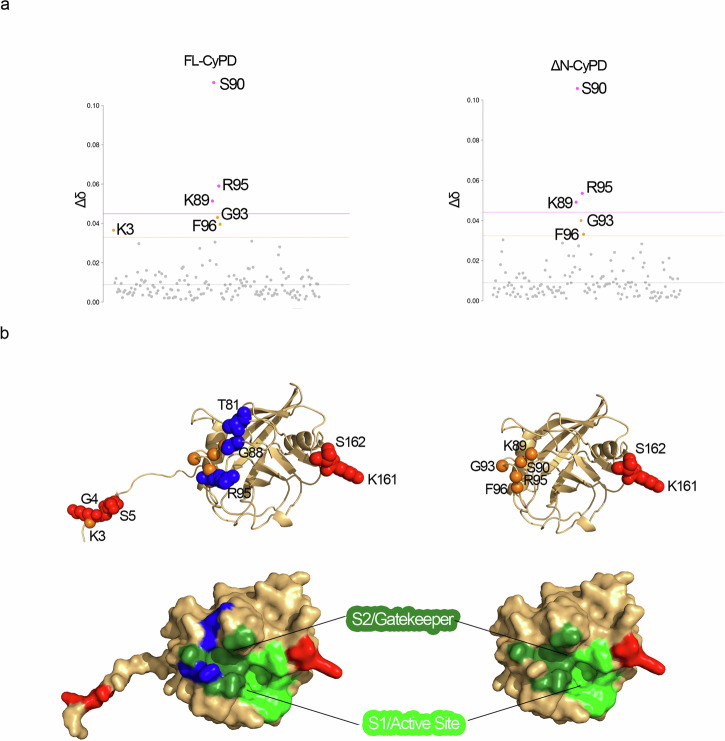
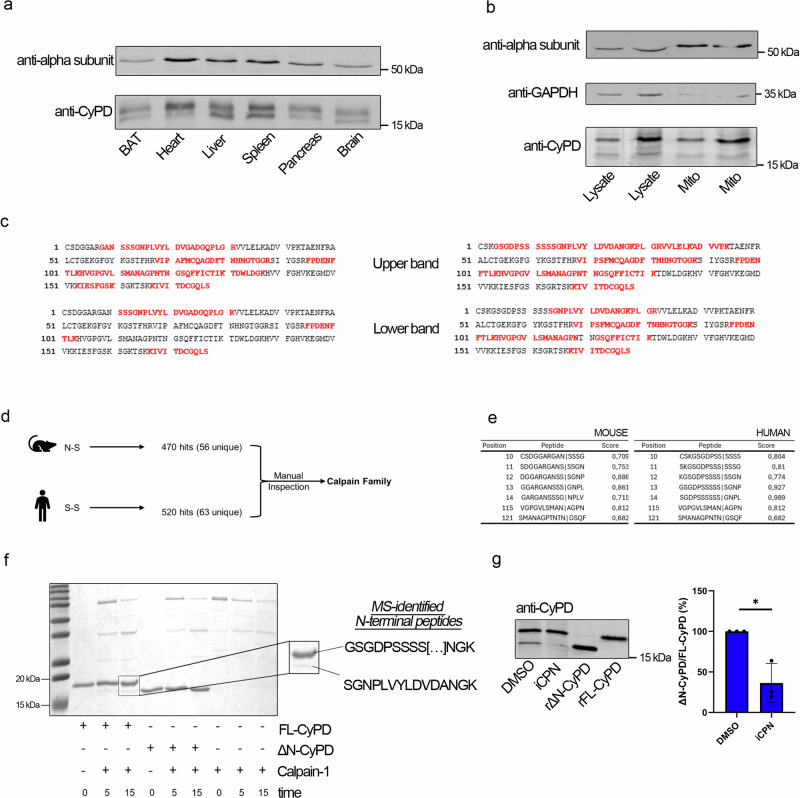
Cyclophilin (CyP) D is a regulator of the mitochondrial F-ATP synthase. Here we report the discovery of a form of CyPD lacking the first 10 (mouse) or 13 (human) N-terminal residues (ΔN-CyPD), a protein region with species-specific features. NMR studies on recombinant human full-length CyPD (FL-CyPD) and ΔN-CyPD form revealed that the N-terminus is highly flexible, in contrast with the rigid globular part. We have studied the interactions of FL and ΔN-CyPD with F-ATP synthase at the OSCP subunit, a site where CyPD binding inhibits catalysis and favors the transition of the enzyme complex to the permeability transition pore. At variance from FL-CyPD, ΔN-CyPD binds OSCP in saline media, indicating that the N-terminus substantially decreases the binding affinity for OSCP. We also provide evidence that calpain 1 is responsible for generation of ΔN-CyPD in cells. Altogether, our work suggests the existence of a novel mechanism of modulation of CyPD through cleavage of its N-terminus that may have significant pathophysiological implications.
© 2024. The Author(s).
Conflict of interest statement
Figures
References
-
- Kajitani, K. et al. Crystal structure of human cyclophilin D in complex with its inhibitor, cyclosporin A at 0.96-Å resolution. Proteins70, 1635–1639 (2007). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
