

Inositol pyrophosphate catabolism by three families of phosphatases regulates plant growth and development
- PMID: 39531477
- PMCID: PMC11611267
- DOI: 10.1371/journal.pgen.1011468
Inositol pyrophosphate catabolism by three families of phosphatases regulates plant growth and development
Abstract
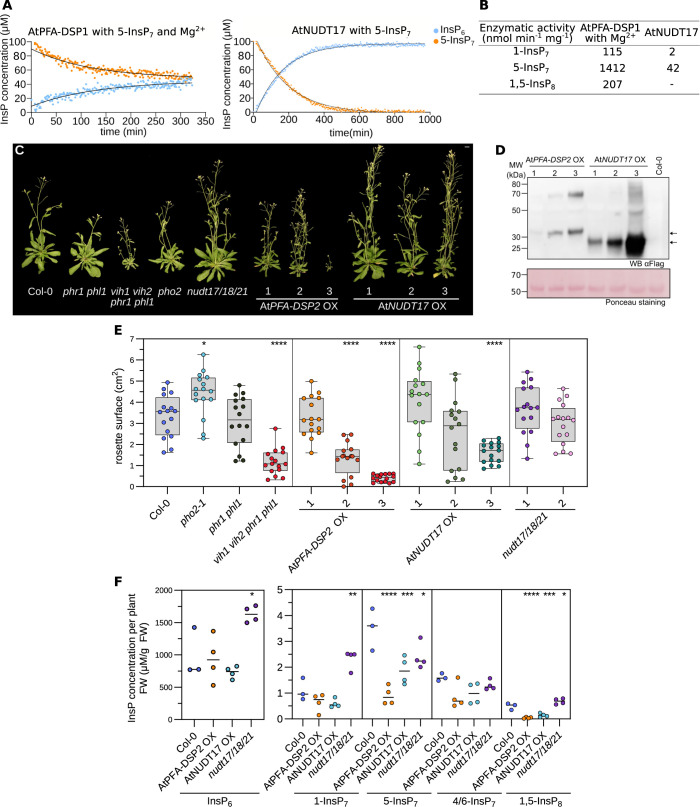
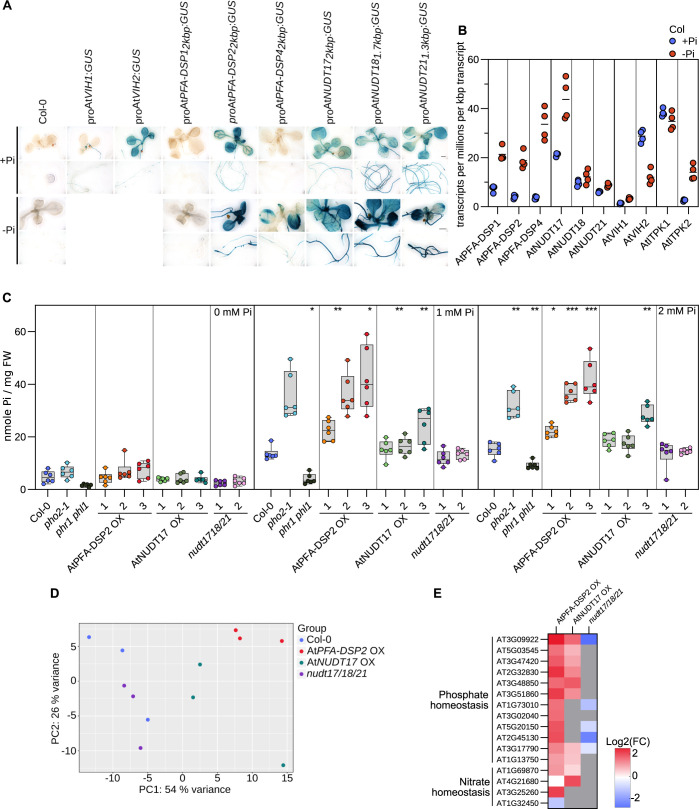
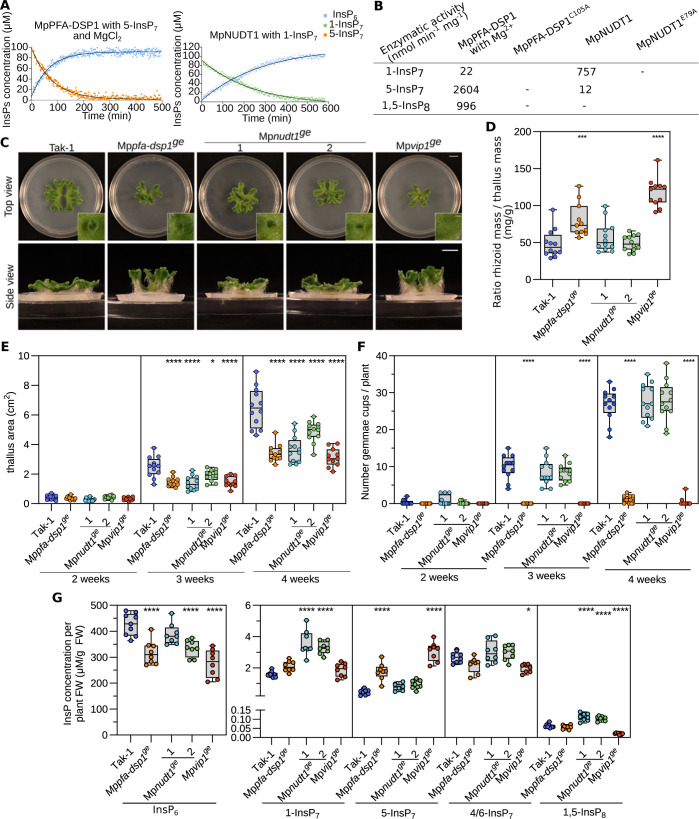
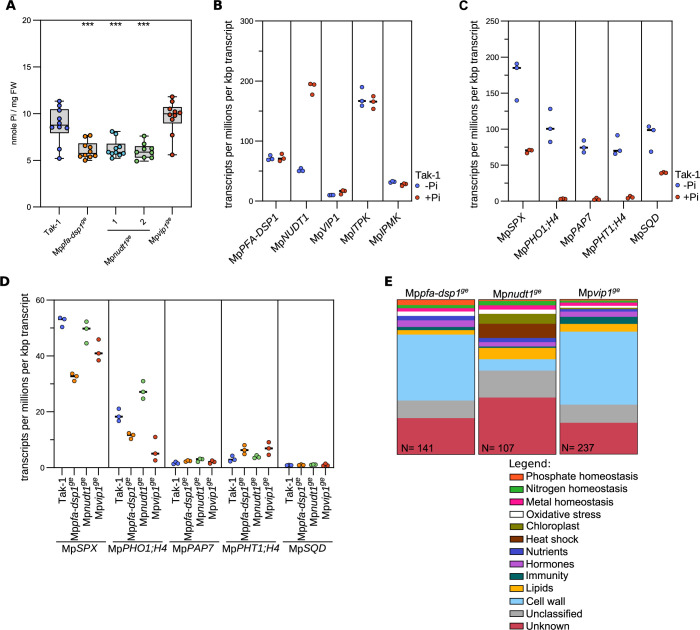
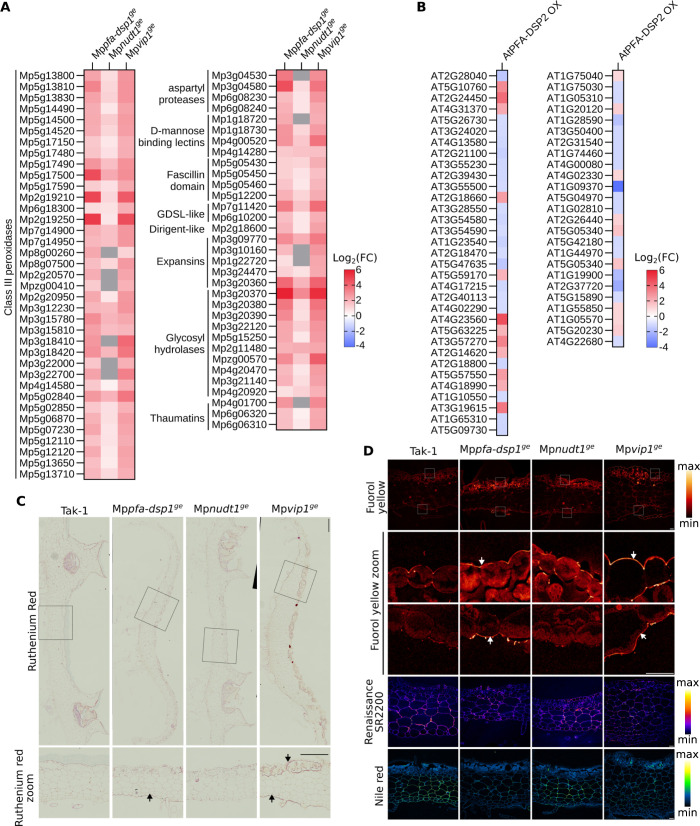
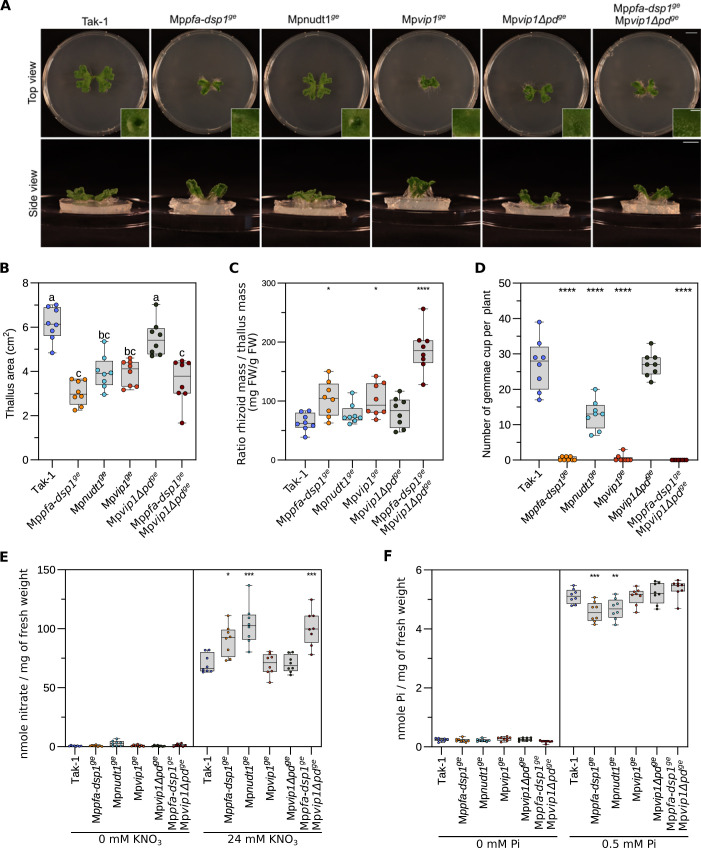
Inositol pyrophosphates (PP-InsPs) are nutrient messengers whose cellular levels are precisely regulated. Diphosphoinositol pentakisphosphate kinases (PPIP5Ks) generate the active signaling molecule 1,5-InsP8. PPIP5Ks harbor phosphatase domains that hydrolyze PP-InsPs. Plant and Fungi Atypical Dual Specificity Phosphatases (PFA-DSPs) and NUDIX phosphatases (NUDTs) are also involved in PP-InsP degradation. Here, we analyze the relative contributions of the three different phosphatase families to plant PP-InsP catabolism. We report the biochemical characterization of inositol pyrophosphate phosphatases from Arabidopsis and Marchantia polymorpha. Overexpression of different PFA-DSP and NUDT enzymes affects PP-InsP levels and leads to stunted growth phenotypes in Arabidopsis. nudt17/18/21 knock-out mutants have altered PP-InsP pools and gene expression patterns, but no apparent growth defects. In contrast, Marchantia polymorpha Mppfa-dsp1ge, Mpnudt1ge and Mpvip1ge mutants display severe growth and developmental phenotypes and associated changes in cellular PP-InsP levels. Analysis of Mppfa-dsp1geand Mpvip1ge mutants supports a role for PP-InsPs in Marchantia phosphate signaling, and additional functions in nitrate homeostasis and cell wall biogenesis. Simultaneous elimination of two phosphatase activities enhanced the observed growth phenotypes. Taken together, PPIP5K, PFA-DSP and NUDT inositol pyrophosphate phosphatases regulate growth and development by collectively shaping plant PP-InsP pools.
Copyright: © 2024 Laurent et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
