

LARP1 binds ribosomes and TOP mRNAs in repressed complexes
- PMID: 39533057
- PMCID: PMC11649897
- DOI: 10.1038/s44318-024-00294-z
LARP1 binds ribosomes and TOP mRNAs in repressed complexes
Abstract
Terminal oligopyrimidine motif-containing mRNAs (TOPs) encode all ribosomal proteins in mammals and are regulated to tune ribosome synthesis to cell state. Previous studies have implicated LARP1 in 40S- or 80S-ribosome complexes that are thought to repress and stabilize TOPs. However, a molecular understanding of how LARP1 and TOPs interact with these ribosome complexes is lacking. Here, we show that LARP1 directly binds non-translating ribosomal subunits. Cryo-EM structures reveal a previously uncharacterized domain of LARP1 bound to and occluding the mRNA channel of the 40S subunit. Increased availability of free ribosomal subunits downstream of various stresses promote 60S joining at the same interface to form LARP1-80S complexes. Simultaneously, LARP1 engages the TOP via its previously characterized La/PAM2 and DM15 domains. Contrary to expectations, ribosome binding within these complexes is not required for LARP1-mediated TOP repression or stabilization, two canonical LARP1 functions. Together, this work provides molecular insight into how LARP1 directly binds ribosomal subunits and challenges existing models describing the function of repressed LARP1-40S/80S-TOP complexes.
Keywords: Cryo-EM; LARP1; Ribosome; TOP mRNA; Translation.
© 2024. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. RG is a member of the Advisory Editorial Board of The EMBO Journal. This has no bearing on the editorial consideration of this article for publication. RG is on the scientific advisory board of Alltrna, Initial Therapeutics, and Arrakis Pharmaceuticals and serves as a consultant for Vertex Pharmaceuticals, Bristol-Myers Squibb (Celgene), Monta Rosa Therapeutics, and Flagship Pioneering. RG previously served on the scientific advisory board at Moderna. WT has two patents (8,748,091 and 8,394,584) licensed to ONT and received reimbursement for travel, accommodation, and/or conference fees to speak at events organized by ONT.
Figures
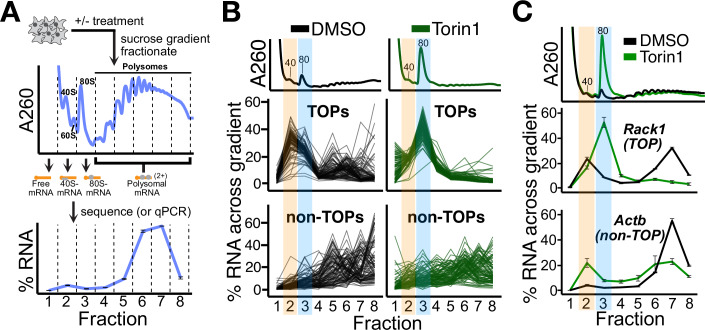




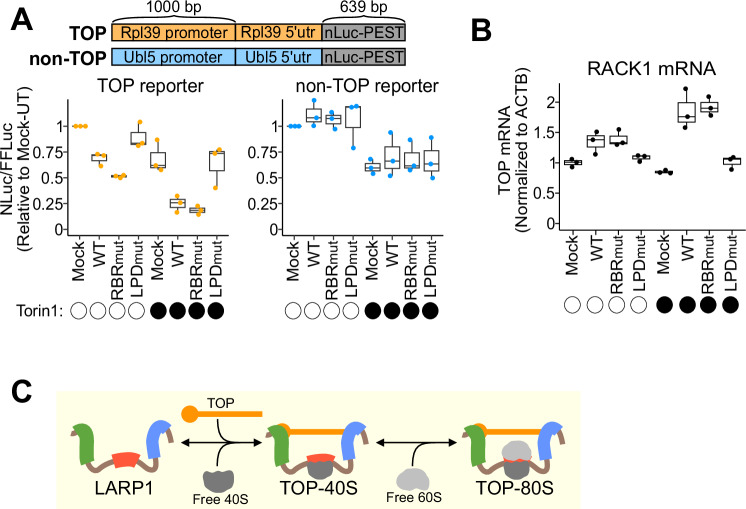
References
-
- Ameismeier M, Cheng J, Berninghausen O, Beckmann R (2018) Visualizing late states of human 40S ribosomal subunit maturation. Nature 558:249–253 - PubMed
-
- Aoki K, Adachi S, Homoto M, Kusano H, Koike K, Natsume T (2013) LARP1 specifically recognizes the 3’ terminus of poly(A) mRNA. FEBS Lett 587:2173–2178 - PubMed
MeSH terms
Substances
Grants and funding
- 22ZR1413600/Shanghai Municipal Science and Technology Commission
- F30 CA260910/CA/NCI NIH HHS/United States
- HG010538/HHS | NIH | National Human Genome Research Institute (NHGRI)
- 2023YFC2413204/National Key R & D Program of China
- 22410712400/Shanghai Municipal Science and Technology Commission
- T32GM136577/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- 32371350/National Natural Science Foundation of China
- F30CA260910/HHS | NIH | National Cancer Institute (NCI)
- T32 GM136577/GM/NIGMS NIH HHS/United States
- HHMI_GREEN/Howard Hughes Medical Institute (HHMI)
- R01 HG010538/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
