

Inserting Omp22 into the flagellin protein, replacing its hypervariable region, results in stronger protection against lethal Acinetobacter baumannii infection
- PMID: 39533090
- PMCID: PMC11557591
- DOI: 10.1038/s41598-024-79013-x
Inserting Omp22 into the flagellin protein, replacing its hypervariable region, results in stronger protection against lethal Acinetobacter baumannii infection
Abstract
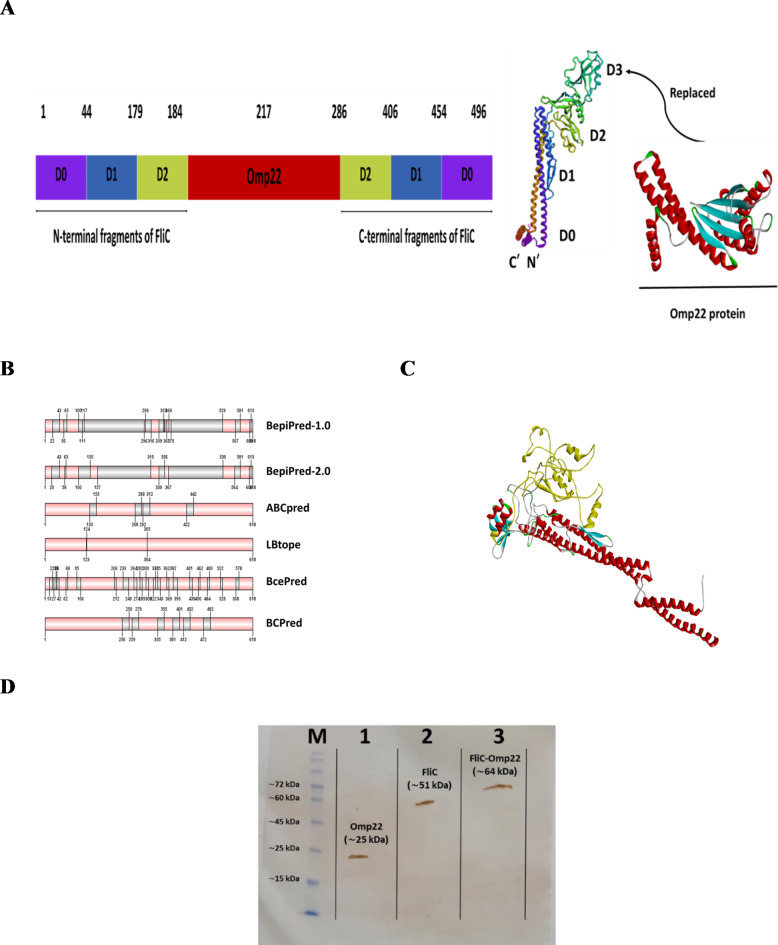
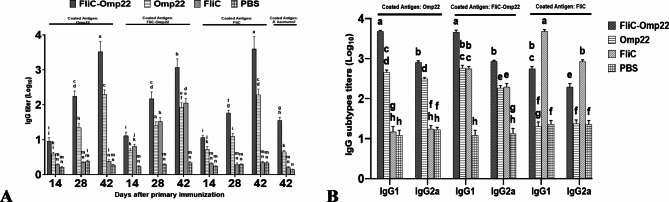
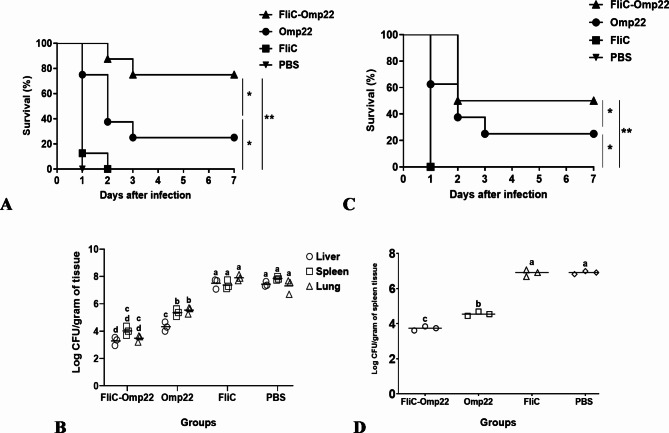
Acinetobacter baumannii, a common nosocomial pathogen, is known for its rapid acquisition of antimicrobial resistance, underscoring the urgent need to develop an effective vaccine against this pathogen. Outer membrane protein 22 (Omp22) regulates the biogenesis of outer membrane vesicles to transport virulence-promoting factors into the host cells and facilitates the progression of A. baumannii infection. In this study, we used a mouse model to assess a vaccine's immunogenicity and protective efficacy using recombinant Omp22 protein within the hypervariable region of flagellin (FliC-Omp22). FliC-Omp22 demonstrated superior protection following challenge with a lethal dose of multidrug-resistant (MDR) A. baumannii strain 58ST compared to Omp22 alone. In addition, it elicited increased IgG1/IgG2a and IL-4/IFN-γ ratios, indicating a predominant Th2 immune response. Furthermore, the FliC-Omp22 vaccination elicited strong specific antibodies that inhibited the adhesion and invasion of A. baumannii 58ST and enhanced the opsonic killing activity against the pathogen. FliC-Omp22 immunization significantly reduced bacterial loads in infected mice's spleen, lungs, and liver, thereby improving their survival against the lethal infection caused by MDR A. baumannii 58ST. This study suggests that integrating Omp22 into the hypervariable domain of flagellin holds promise for developing an effective vaccine against A. baumannii infections.
Keywords: Acinetobacter baumannii.; Flagellin.; FliC.; Immunization.; Omp22.; Sepsis.
© 2024. The Author(s).
Conflict of interest statement
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
