

TRIM28 regulates the coagulation cascade inhibited by p72 of African swine fever virus
- PMID: 39533356
- PMCID: PMC11559047
- DOI: 10.1186/s13567-024-01407-6
TRIM28 regulates the coagulation cascade inhibited by p72 of African swine fever virus
Abstract
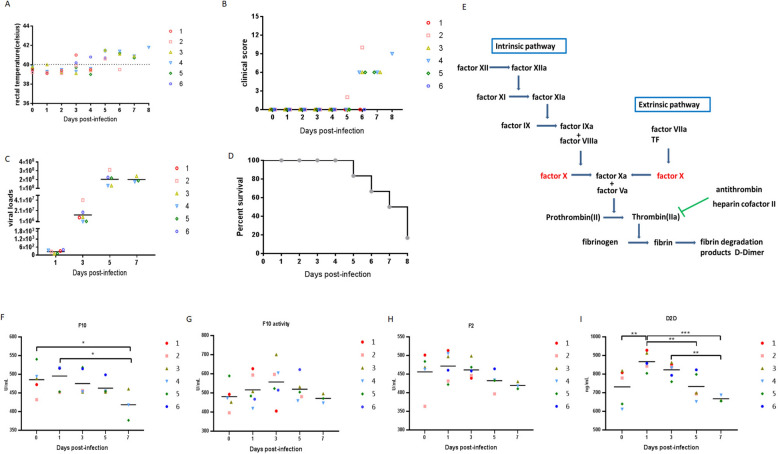
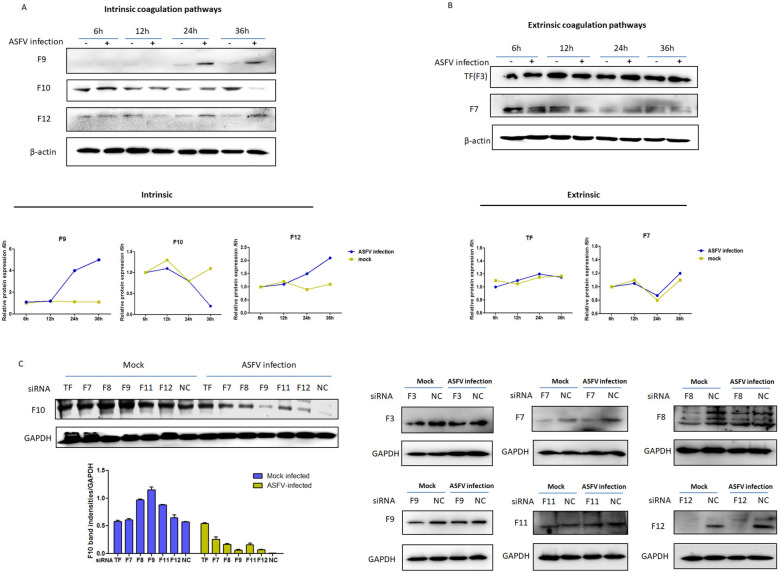
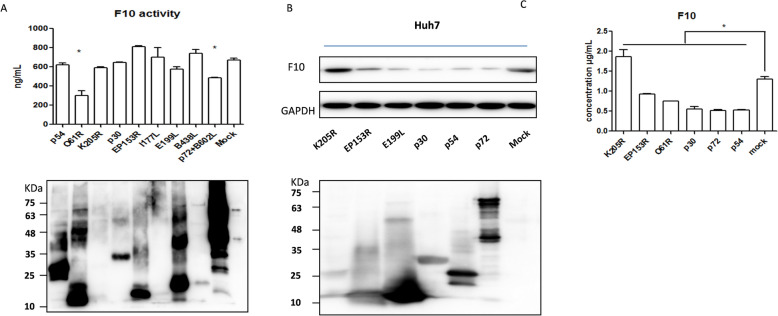
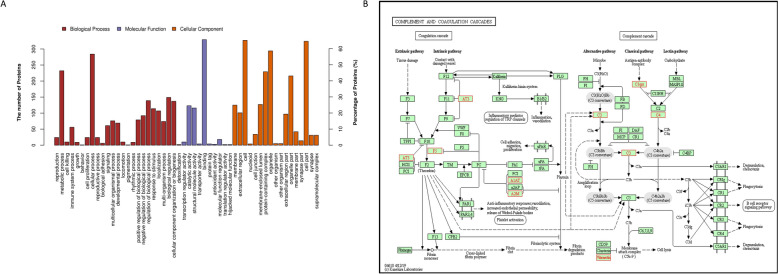
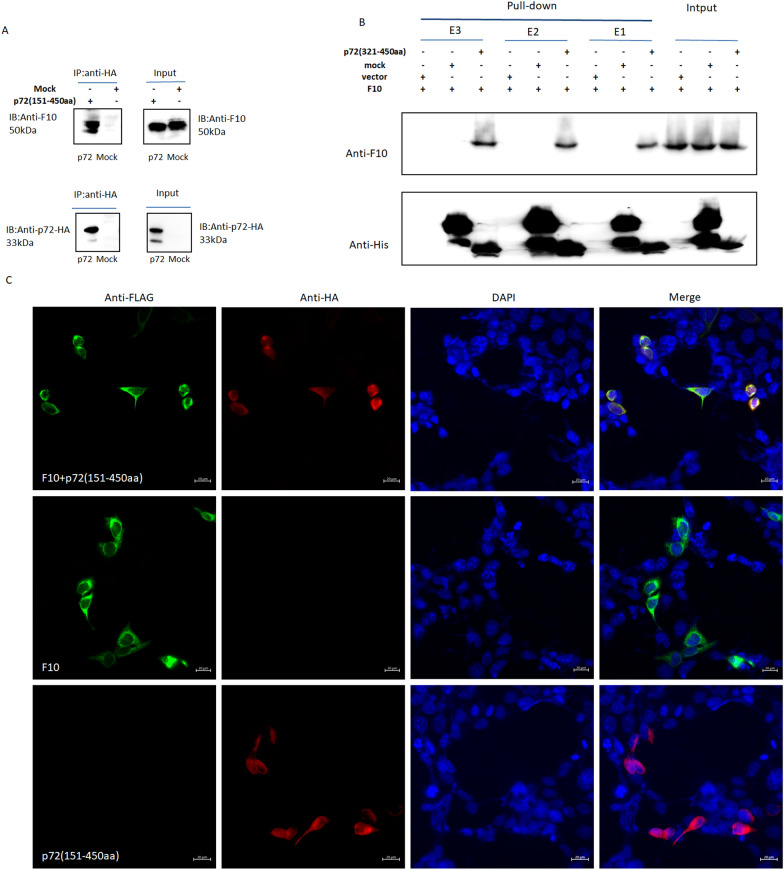
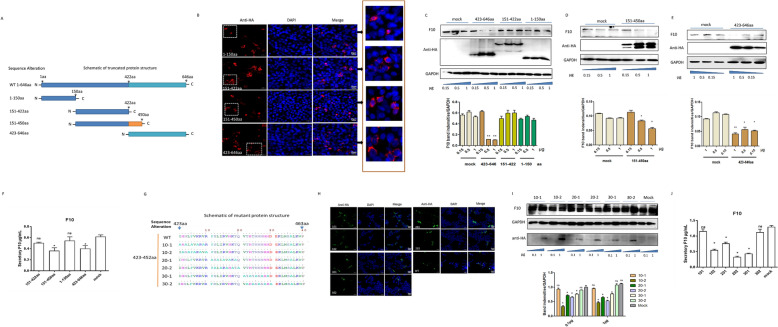
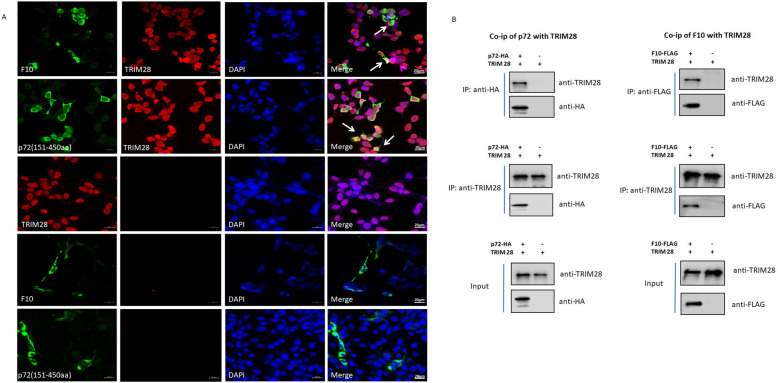
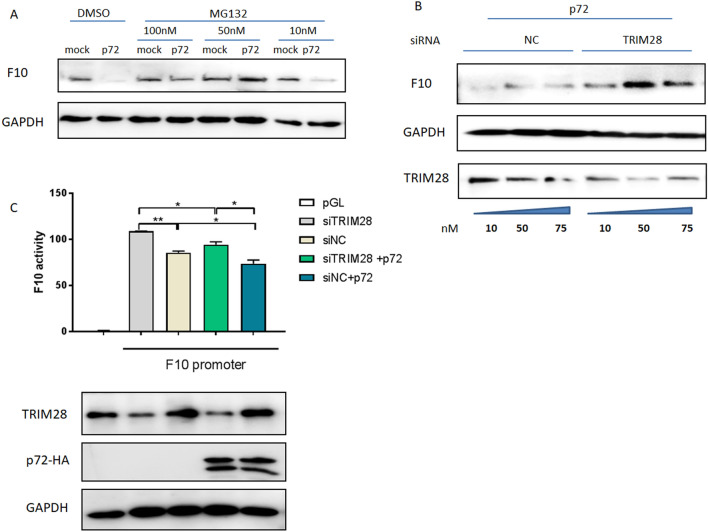
In 2018, African swine fever virus (ASFV) emerged in China, causing extremely serious economic losses to the domestic pig industry. Infection with ASFV can cause disseminated coagulation, leading to the consumption of platelets and coagulation factors and severe bleeding. However, the mechanism of virus-induced coagulation has yet to be established. In our study, ASFV downregulated the coagulation process, as detected by D-dimer (D2D) and Factor X (F10) expression in pigs challenged with ASFV HLJ/18. In vitro, ASFV infection increased Factor IX (F9) and Factor XII (F12) expression while downregulating F10 expression in porcine alveolar macrophages (PAMs). African swine fever virus induced both intrinsic and extrinsic coagulation cascades. In addition, several encoded proteins affect the expression of the crucial coagulation protein F10, and among the encoded proteins, p72 inhibits the activity and expression of F10. Proteomic analysis also revealed that p72 is involved in the coagulation cascade. p72 can interact with F10, and its inhibitory functional domains include amino acids 423-432 and amino acids 443-452. Finally, we found that F10 and p72 interact with tripartite motif-containing protein 28 (TRIM28). TRIM28 knockdown resulted in a decrease in F10 expression. Importantly, TRIM28 contributes to the reduction in F10 protein expression regulated by p72. Our findings revealed an inhibitory effect of the viral protein p72 on the ASFV infection-induced coagulation cascade and revealed a role of TRIM28 in reducing F10 expression, revealing a molecular mechanism of ASFV-associated coagulation.
Keywords: ASFV; P72; TRIM28; coagulation cascade; factor 10.
© 2024. The Author(s).
Conflict of interest statement
Figures
Similar articles
-
Complete Genome Sequencing and Comparative Phylogenomics of Nine African Swine Fever Virus (ASFV) Isolates of the Virulent East African p72 Genotype IX without Viral Sequence Enrichment.Viruses. 2024 Sep 14;16(9):1466. doi: 10.3390/v16091466. Viruses. 2024. PMID: 39339942 Free PMC article.
-
Autophagy promotes p72 degradation and capsid disassembly during the early phase of African swine fever virus infection.J Virol. 2025 Jan 31;99(1):e0170124. doi: 10.1128/jvi.01701-24. Epub 2024 Dec 17. J Virol. 2025. PMID: 39688418 Free PMC article.
-
African Swine Fever Virus E120R Protein Inhibits Interferon Beta Production by Interacting with IRF3 To Block Its Activation.J Virol. 2021 Aug 25;95(18):e0082421. doi: 10.1128/JVI.00824-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34190598 Free PMC article.
-
Research progress on the proteins involved in African swine fever virus infection and replication.Front Immunol. 2022 Jul 22;13:947180. doi: 10.3389/fimmu.2022.947180. eCollection 2022. Front Immunol. 2022. PMID: 35935977 Free PMC article. Review.
-
Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review.Front Immunol. 2021 Sep 6;12:715582. doi: 10.3389/fimmu.2021.715582. eCollection 2021. Front Immunol. 2021. PMID: 34552586 Free PMC article. Review.
References
-
- Dixon LK, Sun H, Roberts H (2019) African swine fever. Antiviral Res 165:34–41 - PubMed
-
- Zhou X, Li N, Luo Y, Liu Y, Miao F, Chen T, Zhang S, Cao P, Li X, Tian K, Qiu HJ, Hu R (2018) Emergence of African swine fever in China, 2018. Transbound Emerg Dis 65:1482–1484 - PubMed
-
- You SB, Liu TY, Zhang M, Zhao X, Dong YZ, Wu B, Wang YZ, Li J, Wei XJ, Shi BF (2021) African swine fever outbreaks in China led to gross domestic product and economic losses. Nat Food 2:802–808 - PubMed
-
- Salas ML, Andres G (2013) African swine fever virus morphogenesis. Virus Res 173:29–41 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
