

LiaR-dependent gene expression contributes to antimicrobial responses in group A Streptococcus
- PMID: 39535201
- PMCID: PMC11619527
- DOI: 10.1128/aac.00496-24
LiaR-dependent gene expression contributes to antimicrobial responses in group A Streptococcus
Abstract
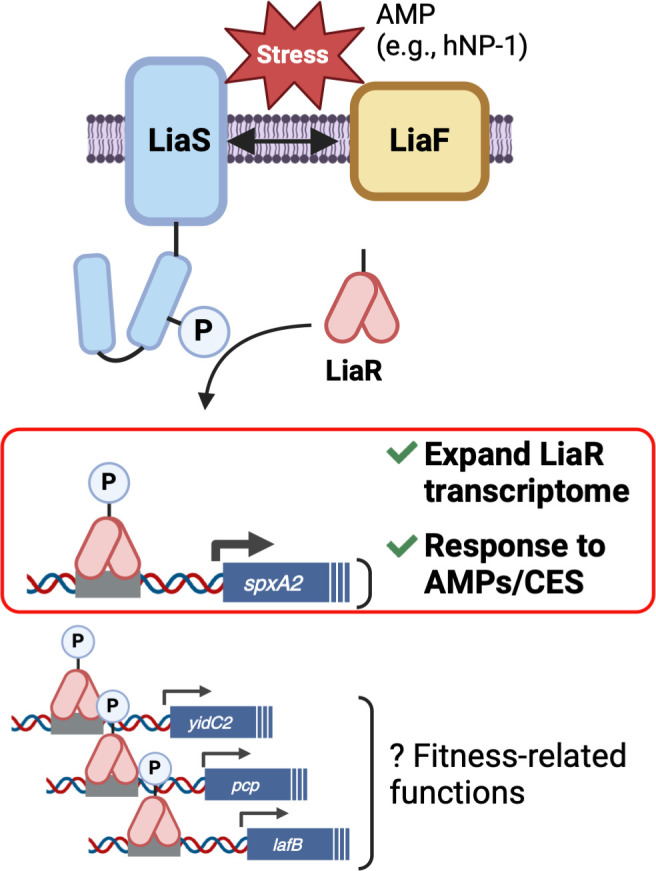
The ability to sense and respond to host defenses is essential for pathogen survival. Some mechanisms involve two-component systems (TCSs) that respond to host molecules, such as antimicrobial peptides (AMPs), and activate specific gene regulatory pathways to aid in survival. Alongside TCSs, bacteria coordinate cell division proteins, chaperones, cell wall sortases, and secretory translocons at discrete locations within the cytoplasmic membrane, referred to as functional membrane microdomains (FMMs). In group A Streptococcus (GAS), the FMM or "ExPortal" coordinates protein secretion, cell wall synthesis, and sensing of AMP-mediated cell envelope stress via the LiaFSR three-component system. Previously, we showed that GAS exposure to a subset of AMPs (α-defensins) activates the LiaFSR system by disrupting LiaF and LiaS co-localization in the ExPortal, leading to increased LiaR phosphorylation, expression of the transcriptional regulator SpxA2, and altered GAS virulence gene expression. The mechanisms by which LiaFSR integrates cell envelope stress with responses to AMP activity and virulence are not fully elucidated. Here, we show the LiaFSR regulon is comprised of genes encoding SpxA2 and three membrane-associated proteins: a PspC domain-containing protein (PCP), the lipoteichoic acid-modifying protein LafB, and the membrane protein insertase YidC2. Our data support that phosphorylated LiaR induces transcription of these genes via a conserved operator, whose disruption attenuates GAS virulence and increases susceptibility to AMPs in a manner primarily dependent on differential expression of SpxA2. Our work expands our understanding of the LiaFSR regulatory network in GAS and identifies targets for further investigation of mechanisms of cell envelope stress tolerance contributing to GAS pathogenesis.
Keywords: LiaFSR; Spx; antimicrobial peptides; gene regulation; global regulatory networks; group A streptococcus; two-component regulatory systems.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Update of
-
LiaR-dependent gene expression contributes to antimicrobial responses in group A Streptococcus.bioRxiv [Preprint]. 2024 Apr 4:2024.04.04.588141. doi: 10.1101/2024.04.04.588141. bioRxiv. 2024. Update in: Antimicrob Agents Chemother. 2024 Dec 5;68(12):e0049624. doi: 10.1128/aac.00496-24. PMID: 38617309 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
