

The impact of inactivation of the GH/IGF axis during aging on healthspan
- PMID: 39535693
- PMCID: PMC12181575
- DOI: 10.1007/s11357-024-01426-3
The impact of inactivation of the GH/IGF axis during aging on healthspan
Abstract
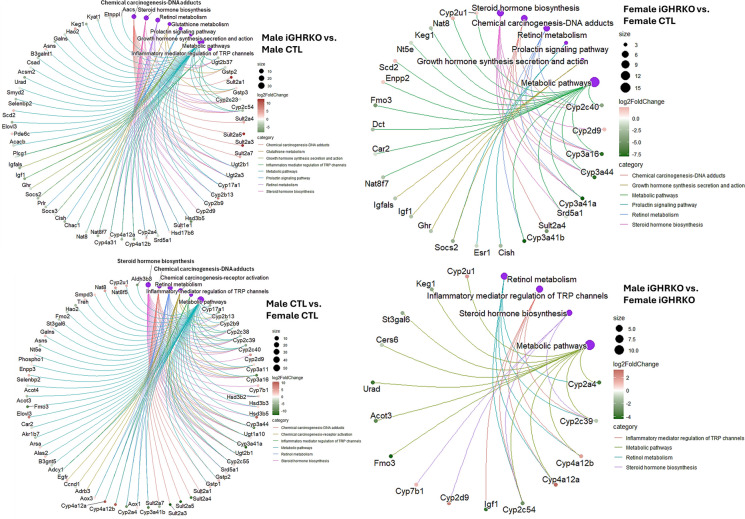
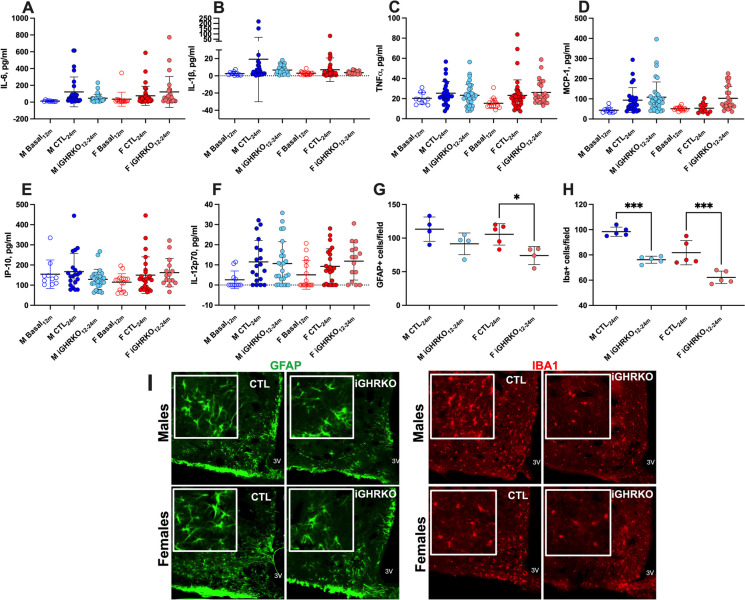
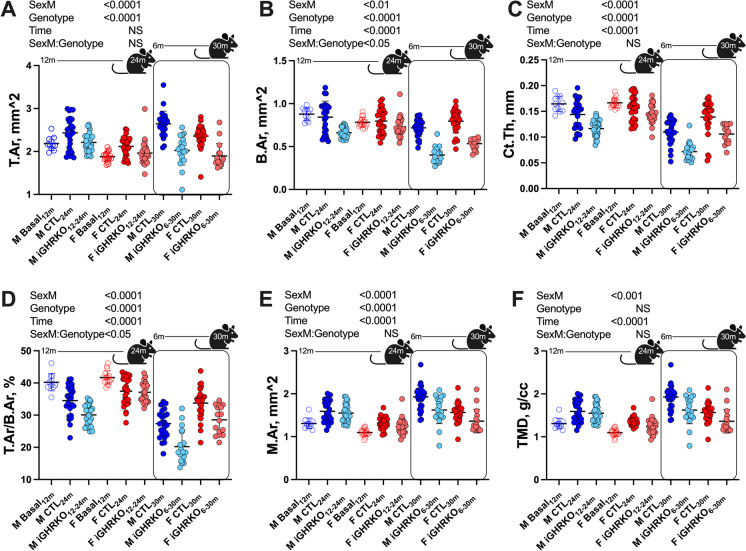
Several mouse lines with congenital growth hormone (GH)/insulin-like growth factor-1 (IGF-1) axis disruption have shown improved health and extended lifespan. The current study investigated how inactivating this axis, specifically during aging, impacts the healthspan. We used a tamoxifen-inducible global GH receptor (GHR) knockout mouse model starting at 12 months and followed the mice until 24 months of age (iGHRKO12-24 mice). We found sex- and tissue-specific effects, with some being pro-aging and others anti-aging. Measuring an array of cytokines in serum revealed that inactivation of the GH/IGF-1 axis at 12 months did not affect systemic inflammation during aging. On the other hand, hypothalamic inflammation was significantly reduced in iGHRKO12-24 mice, evidenced by GFAP+ (glial fibrillary acidic protein, a marker of astrocytes) and Iba-1+ (a marker for microglia). Liver RNAseq analysis indicated feminization of the male transcriptome, with significant changes in the expression of monooxygenase, sulfotransferase, and solute-carrier-transporter gene clusters. Finally, we found impaired bone morphology, more pronounced in male iGHRKO12-24 mice and correlated with GH/IGF-1 inactivation onset age. We conclude that inhibiting the GH/IGF-1 axis during aging only partially preserves the beneficial healthspan effects observed with congenital GH deficiency.
Keywords: Aging; Bone; Growth hormone; Inflammation; Insulin-like growth factor-1; Liver; Micro-CT.
© 2024. The Author(s), under exclusive licence to American Aging Association.
Conflict of interest statement
Declarations. Ethics approval: Animal protocol was reviewed and approved by the Institutional Animal Care and Use Committee of NYU, NY, NY. Conflict of interest: The authors declare no competing interests.
Figures



References
-
- Hage C, Salvatori R. Growth hormone and aging. Endocrinol Metab Clin North Am. 2023;52(2):245–57. - PubMed
-
- Tavares MR, Frazao R, Donato J. Understanding the role of growth hormone in situations of metabolic stress. J Endocrinol. 2023;256(1). 10.1530/JOE-22-0159. - PubMed
-
- Masternak MM, et al. Dwarf mice and aging. Prog Mol Biol Transl Sci. 2018;155:69–83. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
