

TET2 regulates early and late transitions in exhausted CD8+ T cell differentiation and limits CAR T cell function
- PMID: 39536093
- PMCID: PMC11559603
- DOI: 10.1126/sciadv.adp9371
TET2 regulates early and late transitions in exhausted CD8+ T cell differentiation and limits CAR T cell function
Abstract
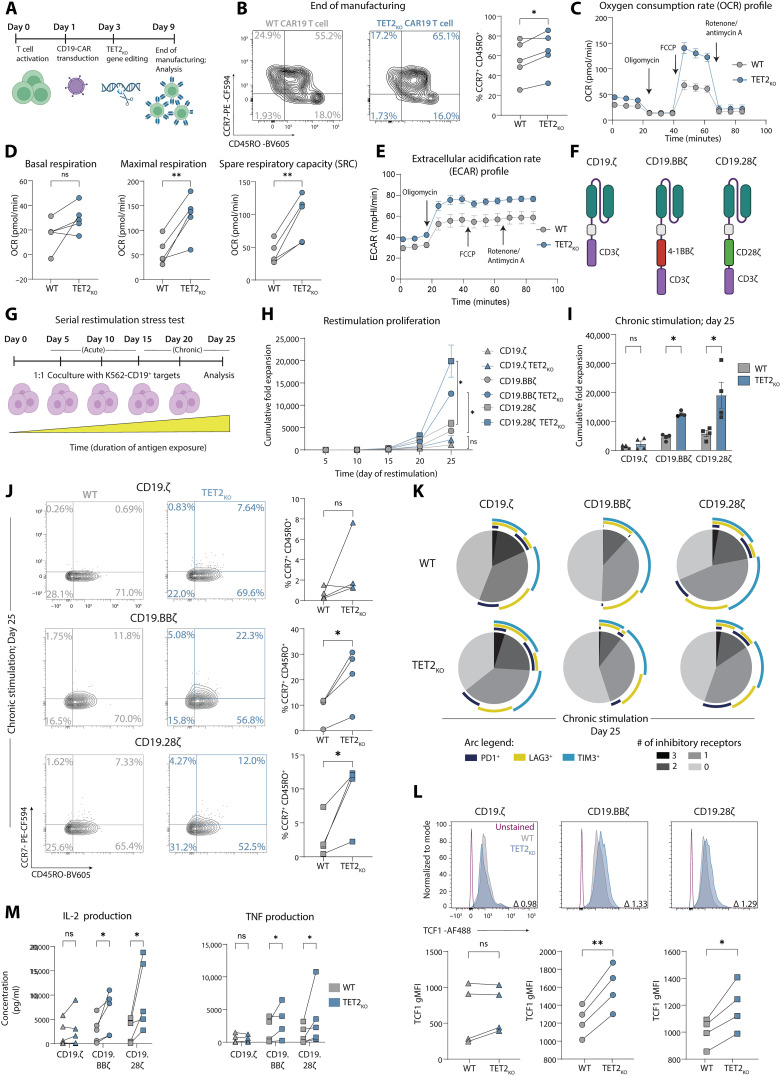
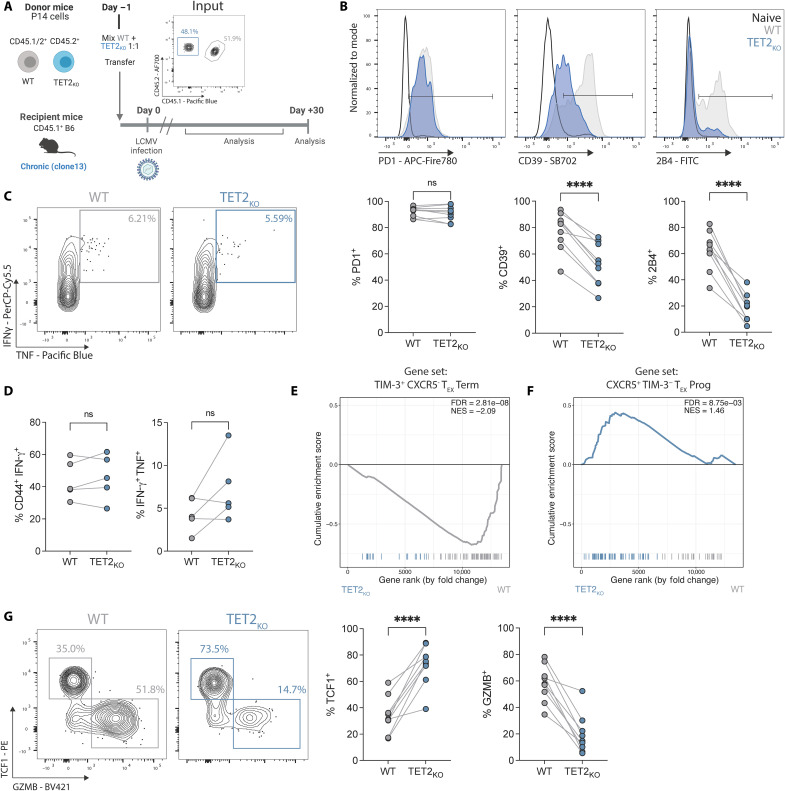
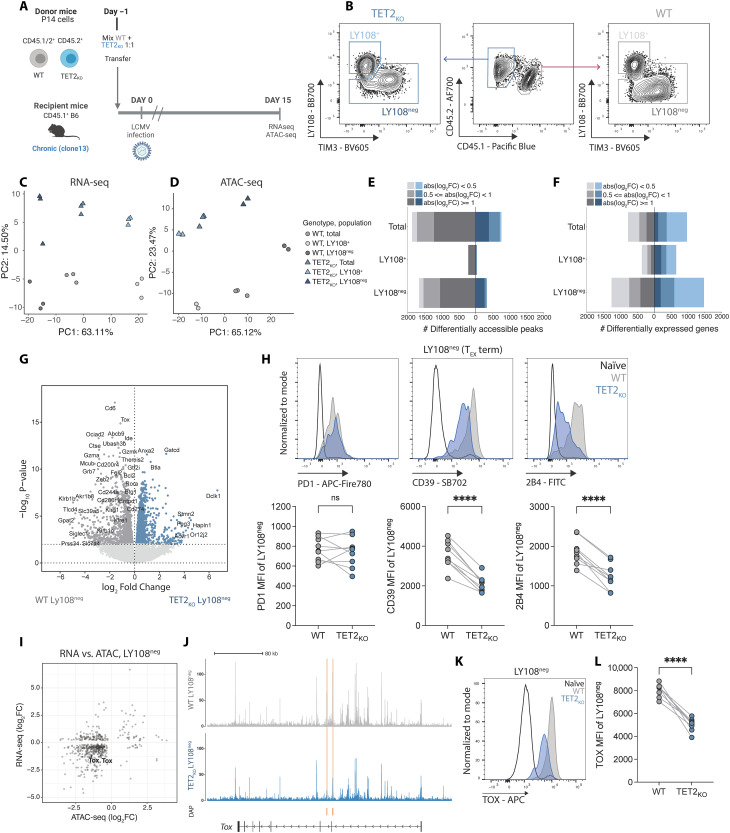
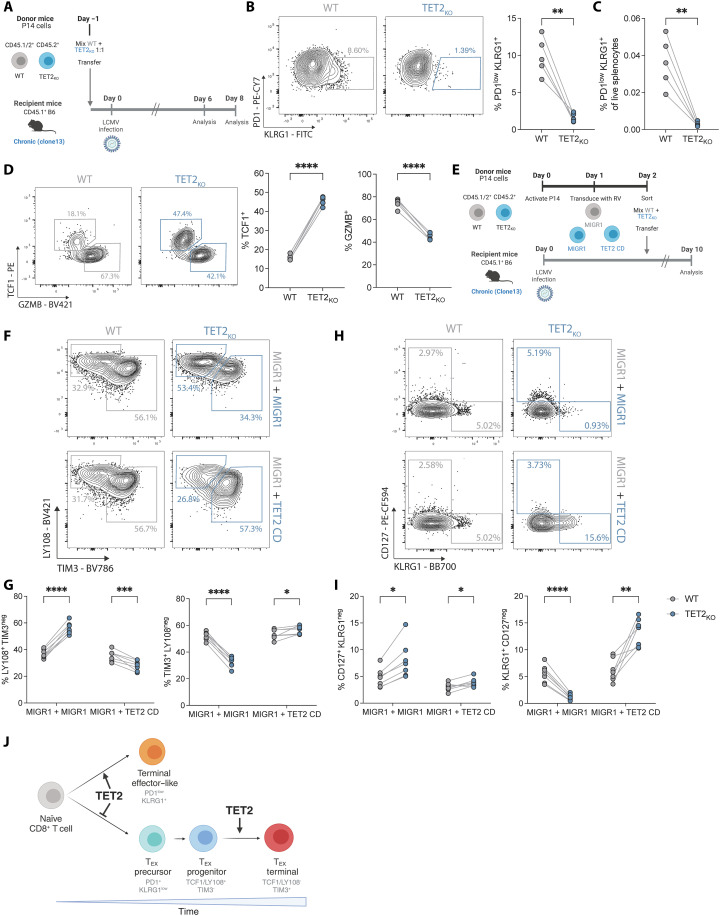
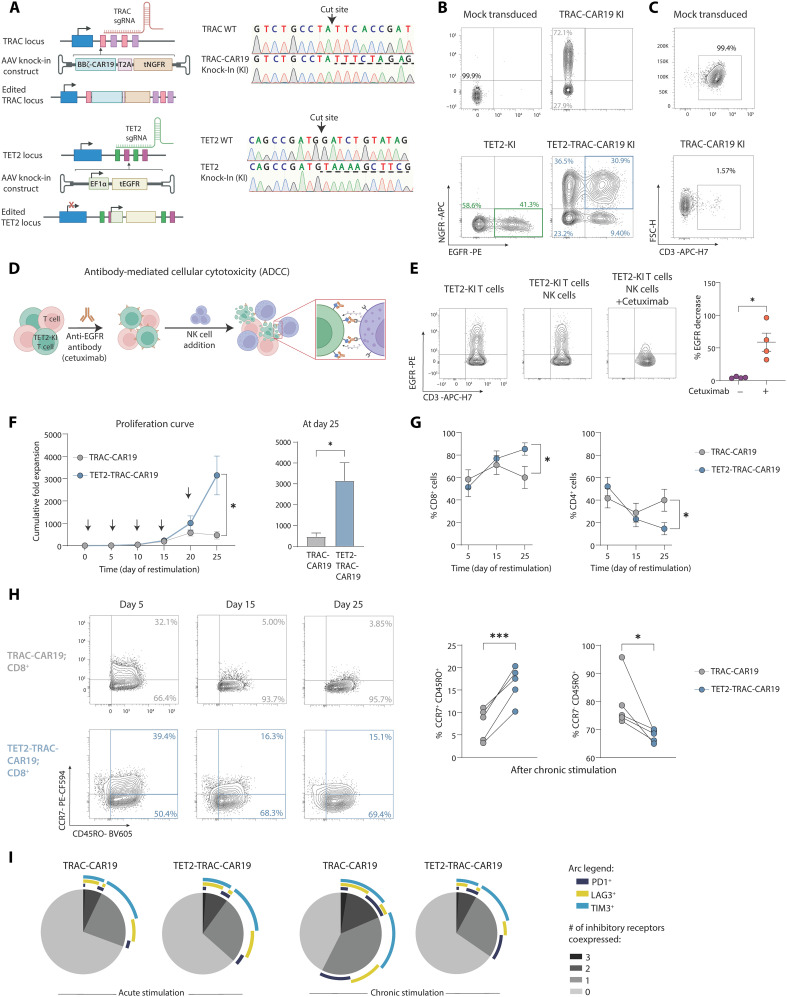
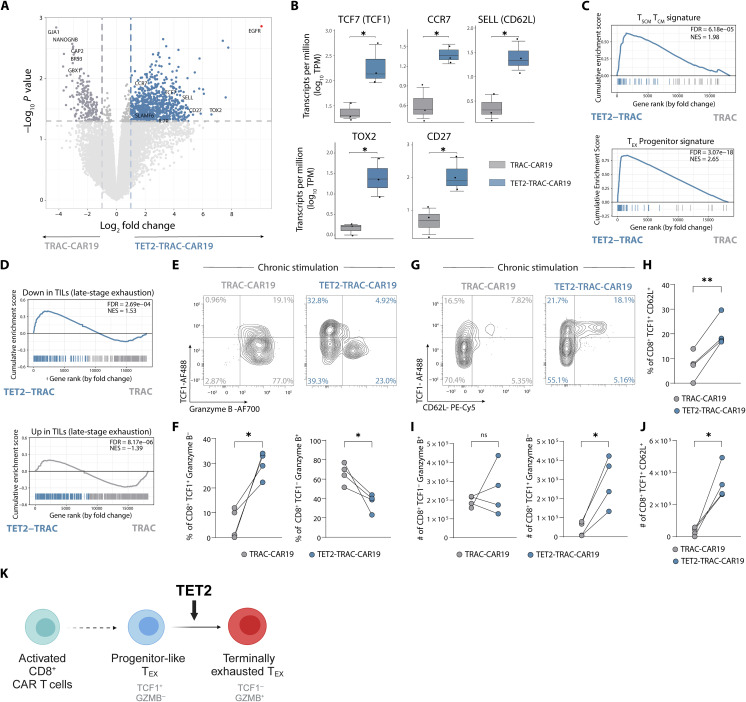
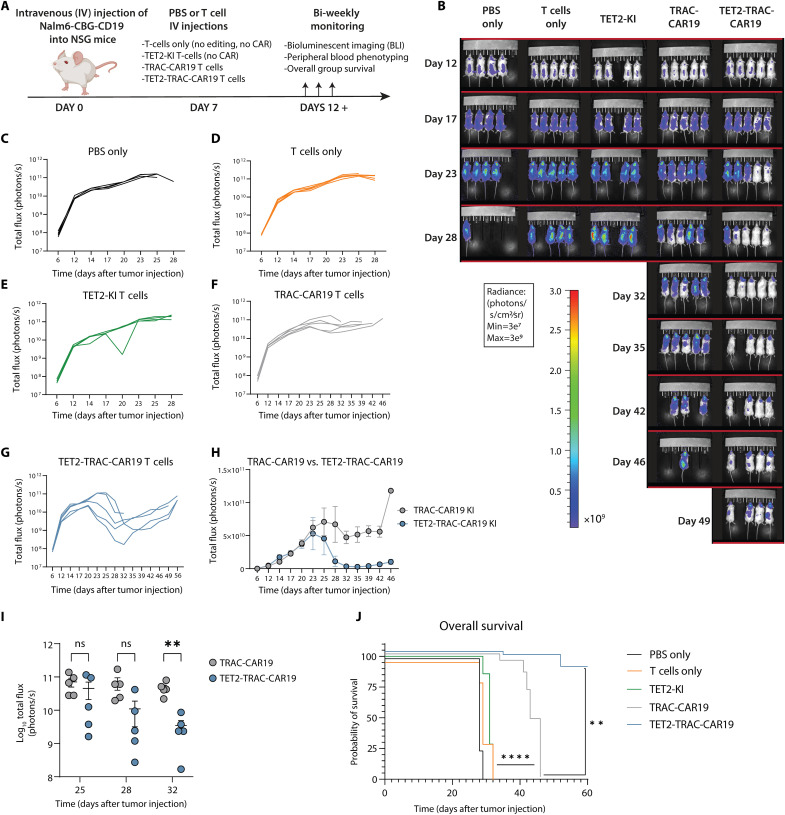
CD8+ T cell exhaustion hampers control of cancer and chronic infections and limits chimeric antigen receptor (CAR) T cell efficacy. Targeting TET2 in CAR T cells provides therapeutic benefit; however, TET2's role in exhausted T cell (TEX) development is unclear. In chronic lymphocytic choriomeningitis virus (LCMV) infection, TET2 drove conversion from stem cell-like TEX progenitors toward terminally differentiated and effector (TEFF)-like TEX. TET2 also enforced a terminally differentiated state in the early bifurcation between TEFF and TEX, indicating broad roles for TET2 in acquisition of effector biology. To exploit the therapeutic potential of TET2, we developed clinically actionable TET2-targeted CAR T cells by disrupting TET2 via knock-in of a safety switch alongside CAR knock-in at the TRAC locus. TET2-targeted CAR T cells exhibited restrained terminal exhaustion in vitro and enhanced antitumor responses in vivo. Thus, TET2 regulates fate transitions in TEX differentiation and can be targeted with a safety mechanism in CAR T cells for improved tumor control.
Figures
References
-
- Blank C. U., Haining W. N., Held W., Hogan P. G., Kallies A., Lugli E., Lynn R. C., Philip M., Rao A., Restifo N. P., Schietinger A., Schumacher T. N., Schwartzberg P. L., Sharpe A. H., Speiser D. E., Wherry E. J., Youngblood B. A., Zehn D., Defining ‘T cell exhaustion’. Nat. Rev. Immunol. 19, 665–674 (2019). - PMC - PubMed
-
- Albelda S. M., CAR T cell therapy for patients with solid tumours: Key lessons to learn and unlearn. Nat. Rev. Clin. Oncol. 21, 47–66 (2024). - PubMed
-
- Lynn R. C., Weber E. W., Sotillo E., Gennert D., Xu P., Good Z., Anbunathan H., Lattin J., Jones R., Tieu V., Nagaraja S., Granja J., de Bourcy C. F. A., Majzner R., Satpathy A. T., Quake S. R., Monje M., Chang H. Y., Mackall C. L., c-Jun overexpression in CAR T cells induces exhaustion resistance. Nature 576, 293–300 (2019). - PMC - PubMed
-
- Long A. H., Haso W. M., Shern J. F., Wanhainen K. M., Murgai M., Ingaramo M., Smith J. P., Walker A. J., Kohler M. E., Venkateshwara V. R., Kaplan R. N., Patterson G. H., Fry T. J., Orentas R. J., Mackall C. L., 4-1BB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptors. Nat. Med. 21, 581–590 (2015). - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
