

Charting and probing the activity of ADARs in human development and cell-fate specification
- PMID: 39537590
- PMCID: PMC11561244
- DOI: 10.1038/s41467-024-53973-0
Charting and probing the activity of ADARs in human development and cell-fate specification
Abstract
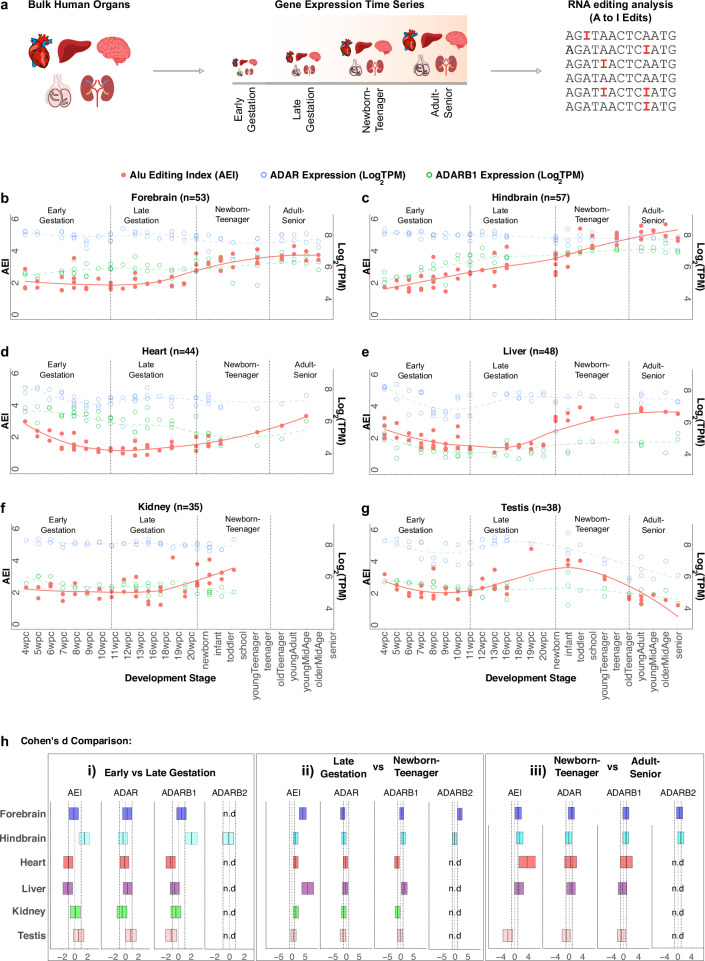
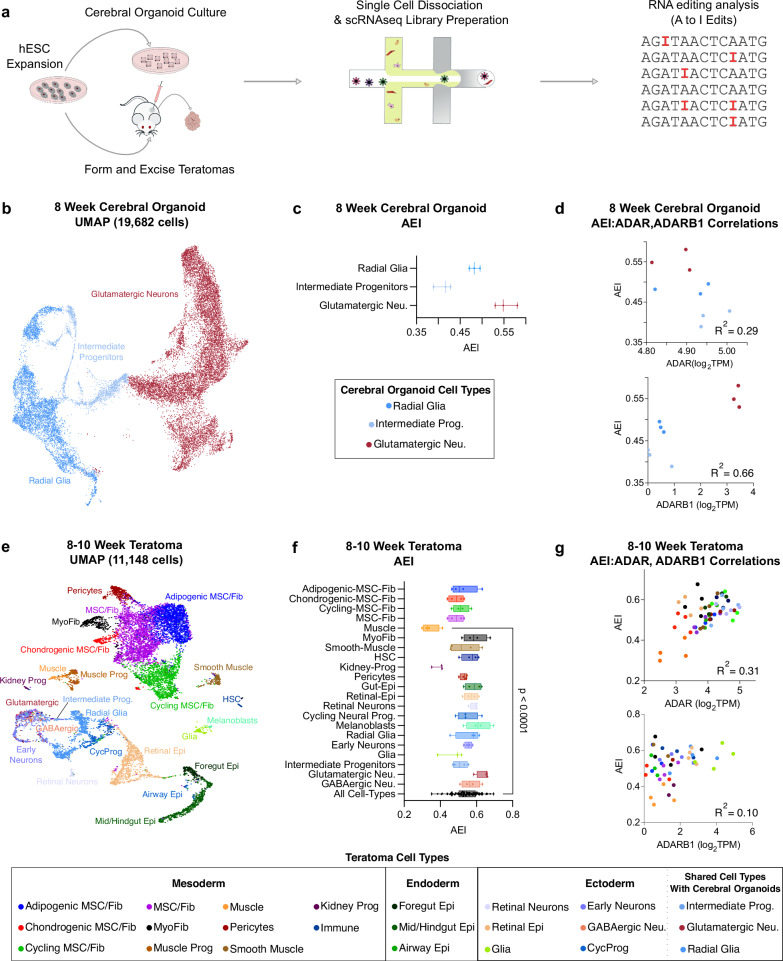
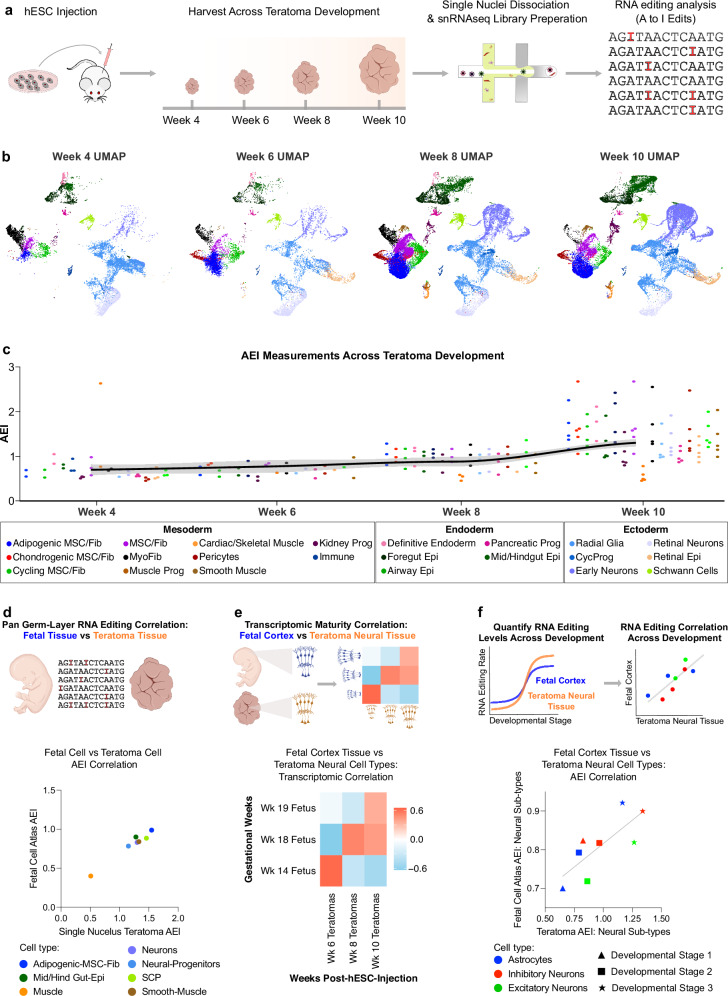
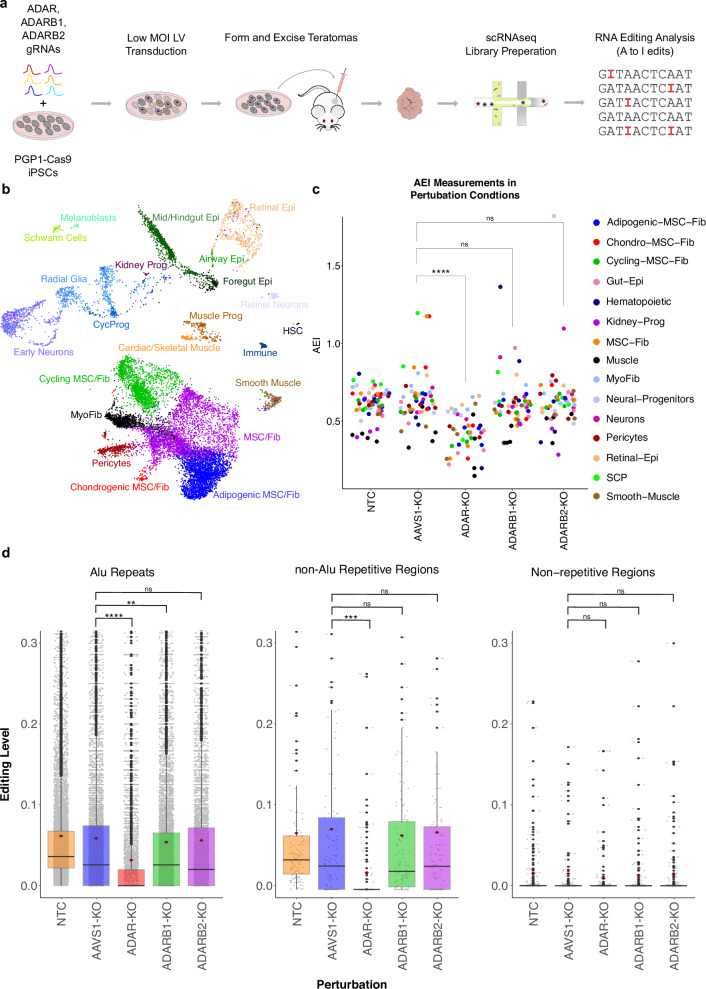
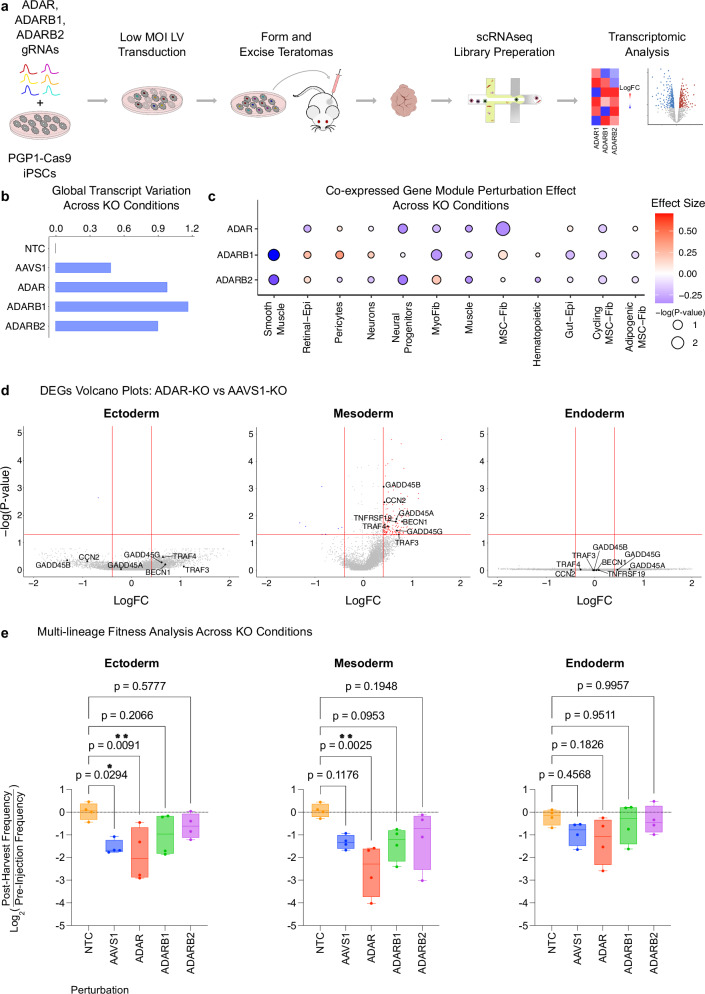
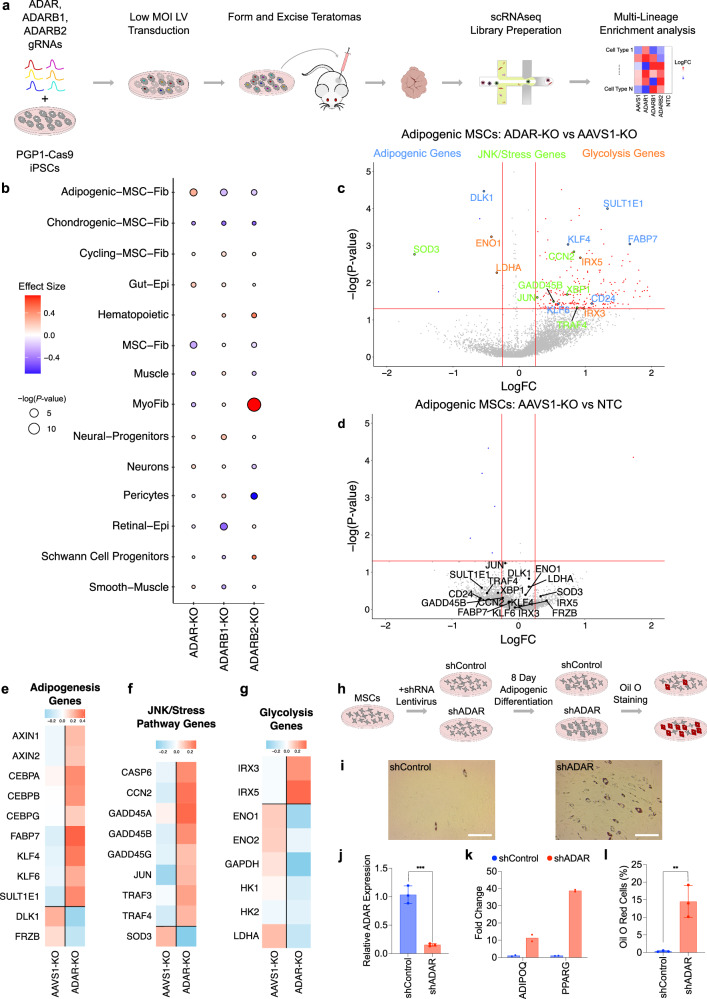
Adenosine deaminases acting on RNA (ADARs) impact diverse cellular processes and pathological conditions, but their functions in early cell-fate specification remain less understood. To gain insights here, we began by charting time-course RNA editing profiles in human organs from fetal to adult stages. Next, we utilized hPSC differentiation to experimentally probe ADARs, harnessing brain organoids as neural specific, and teratomas as pan-tissue developmental models. We show that time-series teratomas faithfully recapitulate fetal developmental trends, and motivated by this, conducted pan-tissue, single-cell CRISPR-KO screens of ADARs in teratomas. Knocking out ADAR leads to a global decrease in RNA editing across all germ-layers. Intriguingly, knocking out ADAR leads to an enrichment of adipogenic cells, revealing a role for ADAR in human adipogenesis. Collectively, we present a multi-pronged framework charting time-resolved RNA editing profiles and coupled ADAR perturbations in developmental models, thereby shedding light on the role of ADARs in cell-fate specification.
© 2024. The Author(s).
Conflict of interest statement
Figures


References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
