

Pck2 association with the plasma membrane and efficient response of the cell integrity pathway require regulation of PI4P homeostasis by exomer
- PMID: 39540318
- PMCID: PMC11561738
- DOI: 10.1098/rsob.240101
Pck2 association with the plasma membrane and efficient response of the cell integrity pathway require regulation of PI4P homeostasis by exomer
Abstract
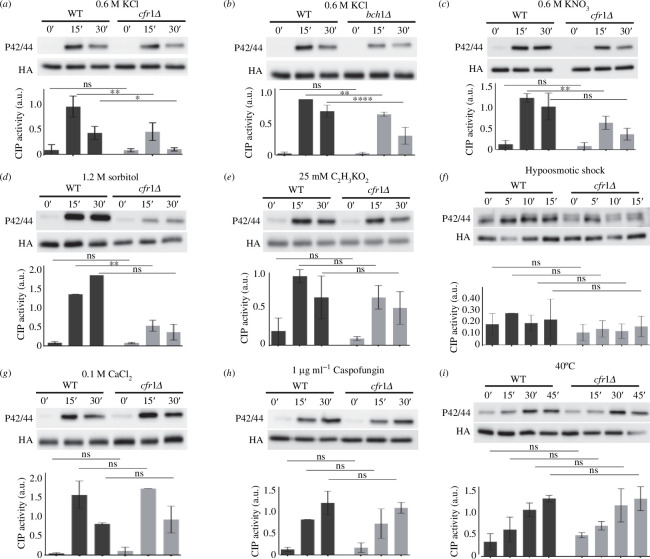
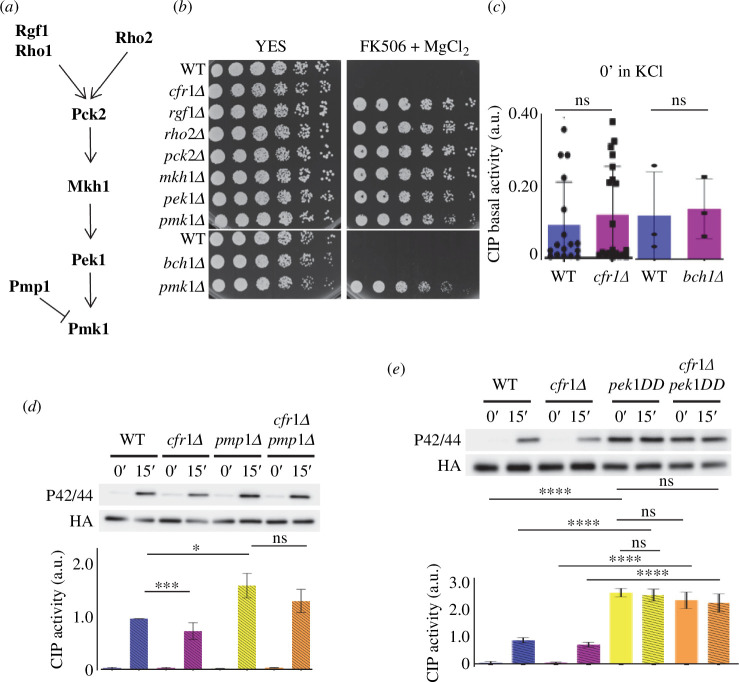
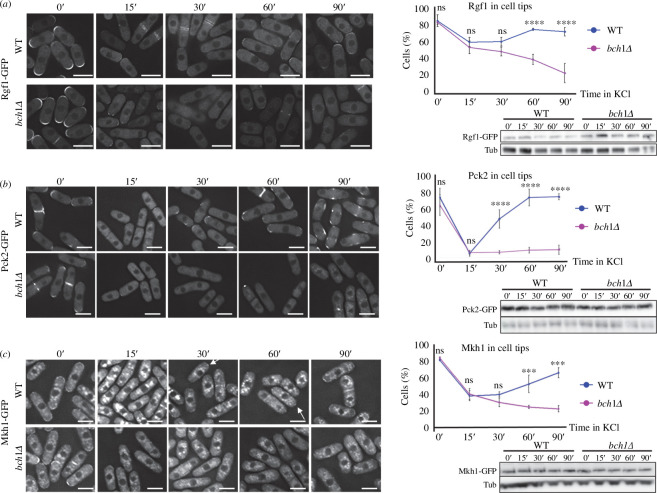
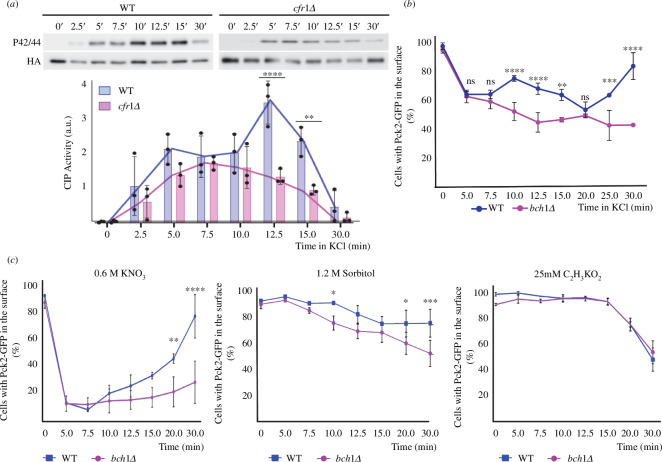
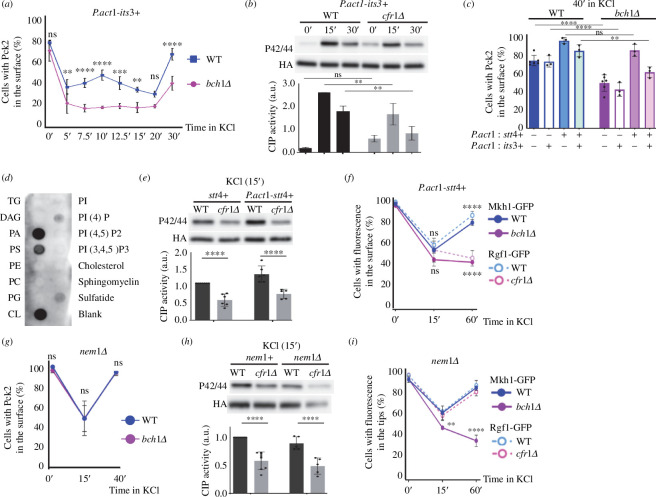
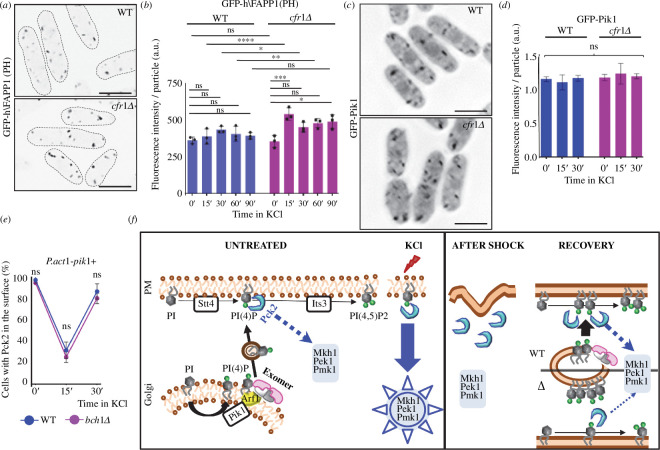
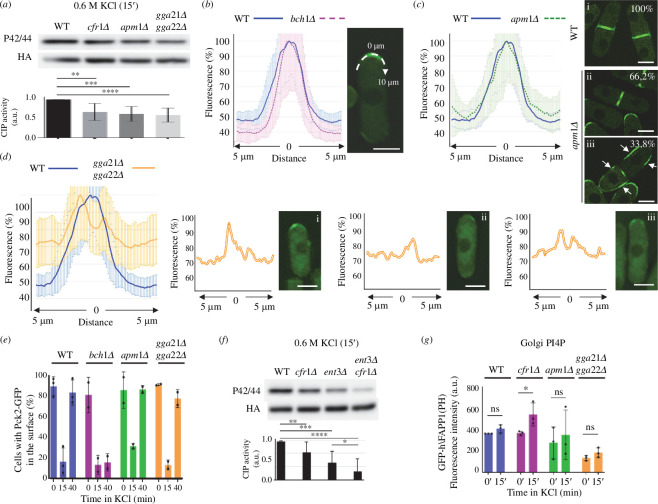
Exomer is a protein complex that facilitates trafficking between the Golgi and the plasma membrane (PM). Schizosaccharomyces pombe exomer is composed of Cfr1 and Bch1, and we have found that full activation of the cell integrity pathway (CIP) in response to osmotic stress requires exomer. In the wild-type, the CIP activators Rgf1 (Rho1 GEF) and Pck2 (PKC homologue) and the MEK kinase Mkh1 localize in the PM, internalize after osmotic shock and re-localize after adaptation. This re-localization is inefficient in exomer mutants. Overexpression of the PM-associated 1-phosphatidylinositol 4-kinase stt4+, and deletion of the nem1+ phosphatase suppress the defects in Pck2 dynamics in exomer mutants, but not their defect in CIP activation, demonstrating that exomer regulates CIP in additional ways. Exomer mutants accumulate PI4P in the TGN, and increasing the expression of the Golgi-associated 1-phosphatidylinositol 4-kinase pik1+ suppresses their defect in Pck2 dynamics. These findings suggest that efficient PI4P transport from the Golgi to the PM requires exomer. Mutants lacking clathrin adaptors are defective in CIP activation, but not in Pck2 dynamics or in PI4P accumulation in the Golgi. Hence, traffic from the Golgi regulates CIP activation, and exomer participates in this regulation through an exclusive mechanism.
Keywords: cell integrity pathway; fission yeast; phosphatidylinositol 4-phosphate; phospholipids; stress response; trans-Golgi network.
Conflict of interest statement
We declare we have no competing interests.
Figures
Similar articles
-
Traffic Through the Trans-Golgi Network and the Endosomal System Requires Collaboration Between Exomer and Clathrin Adaptors in Fission Yeast.Genetics. 2017 Feb;205(2):673-690. doi: 10.1534/genetics.116.193458. Epub 2016 Dec 14. Genetics. 2017. PMID: 27974503 Free PMC article.
-
Exomer Is Part of a Hub Where Polarized Secretion and Ionic Stress Connect.Front Microbiol. 2021 Jul 19;12:708354. doi: 10.3389/fmicb.2021.708354. eCollection 2021. Front Microbiol. 2021. PMID: 34349749 Free PMC article.
-
Rho1 GTPase and PKC ortholog Pck1 are upstream activators of the cell integrity MAPK pathway in fission yeast.PLoS One. 2014 Jan 31;9(1):e88020. doi: 10.1371/journal.pone.0088020. eCollection 2014. PLoS One. 2014. PMID: 24498240 Free PMC article.
-
Regulation and function of PI4P at the Golgi complex.Biochim Biophys Acta Mol Cell Biol Lipids. 2025 Jun;1870(5):159626. doi: 10.1016/j.bbalip.2025.159626. Epub 2025 May 9. Biochim Biophys Acta Mol Cell Biol Lipids. 2025. PMID: 40350028 Review.
-
The Great Escape: how phosphatidylinositol 4-kinases and PI4P promote vesicle exit from the Golgi (and drive cancer).Biochem J. 2019 Aug 28;476(16):2321-2346. doi: 10.1042/BCJ20180622. Biochem J. 2019. PMID: 31462439 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
