

Monocytes give rise to Langerhans cells that preferentially migrate to lymph nodes at steady state
- PMID: 39541348
- PMCID: PMC11588065
- DOI: 10.1073/pnas.2404927121
Monocytes give rise to Langerhans cells that preferentially migrate to lymph nodes at steady state
Abstract
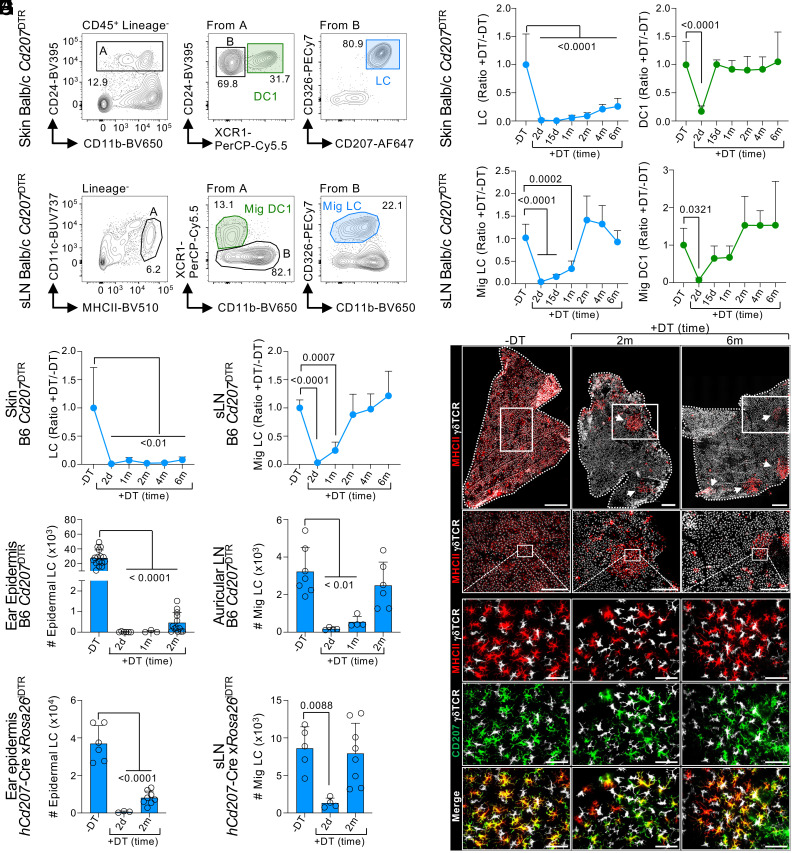
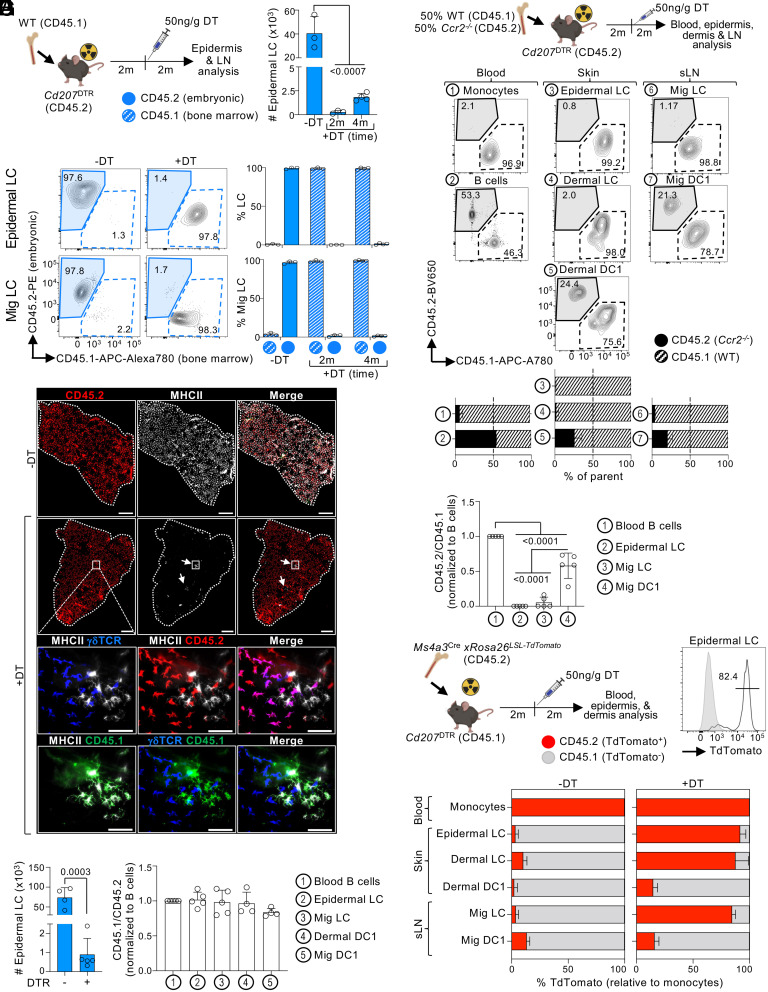
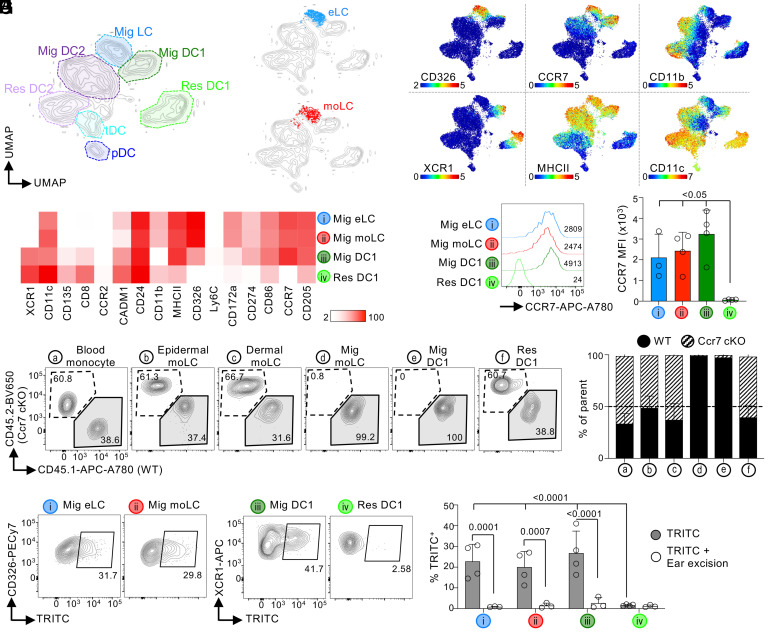
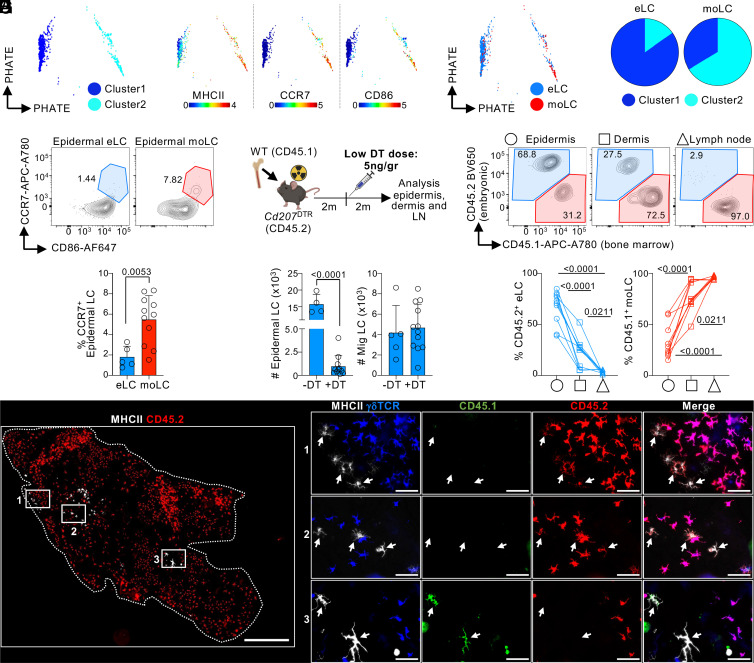
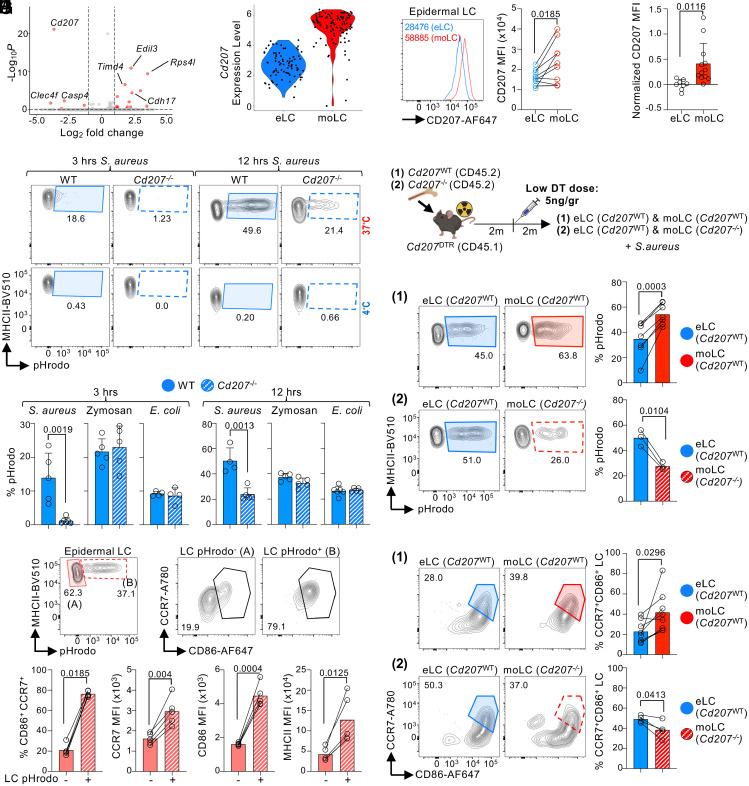
Current evidence suggests that ontogeny may account for the functional heterogeneity of some tissue macrophages, but not others. Here, we asked whether developmental origin drives different functions of skin Langerhans cells (LCs), an embryo-derived mononuclear phagocyte with features of both tissue macrophages and dendritic cells. Using time-course analyses, bone marrow chimeras, and fate tracing models, we found that the complete elimination of embryo-derived LCs at steady state results in their repopulation from circulating monocytes. However, monocyte-derived LCs inefficiently replenished the epidermal niche. Instead, these cells preferentially migrated to skin-draining lymph nodes. Mechanistically, we show that the enhanced migratory capability of monocyte-derived LCs is associated with higher expression of CD207/Langerin, a C-type lectin involved in the capture of skin microbes. Our data demonstrate that ontogeny plays a role in the migratory behavior of epidermal LCs.
Keywords: Langerhans cells; development; macrophages; migration; skin.
Conflict of interest statement
Competing interests statement:J.I. serves on the scientific advisory board of Immunitas Therapeutics. The remaining authors declare no competing interests.
Figures
References
-
- Okabe Y., Medzhitov R., Tissue biology perspective on macrophages. Nat. Immunol. 17, 9–17 (2016). - PubMed
-
- Blériot C., Chakarov S., Ginhoux F., Determinants of resident tissue macrophage identity and function. Immunity 52, 957–970 (2020). - PubMed
-
- Guilliams M., Thierry G. R., Bonnardel J., Bajenoff M., Establishment and maintenance of the macrophage niche. Immunity 52, 434–451 (2020). - PubMed
MeSH terms
Substances
Grants and funding
- AR069953/HHS | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
- AR062595/HHS | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
- CA219994/HHS | NIH | National Cancer Institute (NCI)
- K99 AR062595/AR/NIAMS NIH HHS/United States
- S10 OD016318/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
