

Cap-independent translation directs stress-induced differentiation of the protozoan parasite Toxoplasma gondii
- PMID: 39542243
- PMCID: PMC11697163
- DOI: 10.1016/j.jbc.2024.107979
Cap-independent translation directs stress-induced differentiation of the protozoan parasite Toxoplasma gondii
Abstract
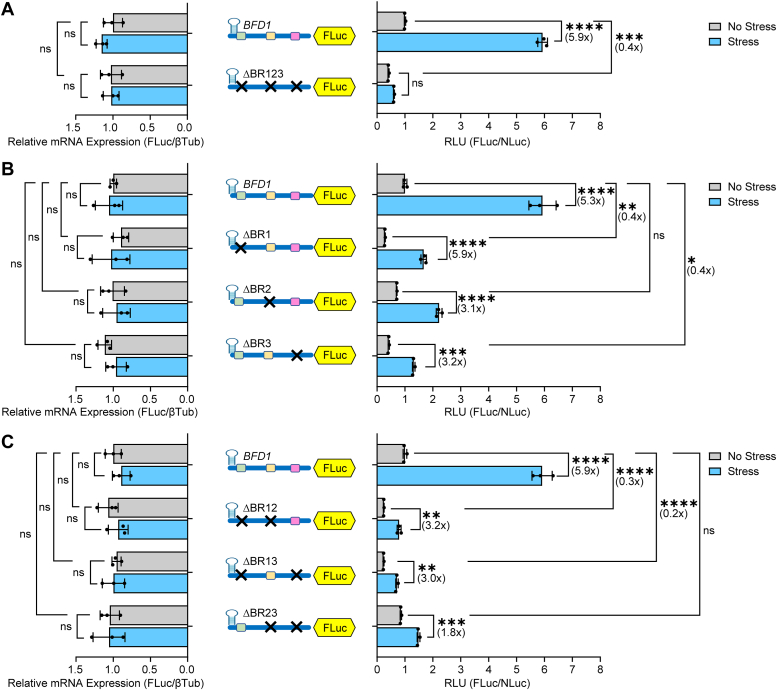
Translational control mechanisms modulate the microbial latency of eukaryotic pathogens, enabling them to evade immunity and drug treatments. The protozoan parasite Toxoplasma gondii persists in hosts by differentiating from proliferative tachyzoites to latent bradyzoites, which are housed inside tissue cysts. Transcriptional changes facilitating bradyzoite conversion are mediated by a Myb domain transcription factor called BFD1, whose mRNA is present in tachyzoites but not translated into protein until stress is applied to induce differentiation. We addressed the mechanisms by which translational control drives BFD1 synthesis in response to stress-induced parasite differentiation. Using biochemical and molecular approaches, we show that the 5'-leader of BFD1 mRNA is sufficient for preferential translation upon stress. The translational control of BFD1 mRNA is maintained when ribosome assembly near its 5'-cap is impaired by insertion of a 5'-proximal stem-loop and upon knockdown of the Toxoplasma cap-binding protein, eIF4E1. Moreover, we determined that a trans-acting RNA-binding protein called BFD2/ROCY1 is necessary for the cap-independent translation of BFD1 through its binding to the 5'-leader. Translation of BFD2 mRNA is also suggested to be preferentially induced under stress but by a cap-dependent mechanism. These results show that translational control and differentiation in Toxoplasma proceed through cap-independent mechanisms in addition to canonical cap-dependent translation. Our identification of cap-independent translation in protozoa underscores the antiquity of this mode of gene regulation in cellular evolution and its central role in stress-induced life-cycle events.
Keywords: Toxoplasma; gene expression; parasite; stress response; translation.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: R. C. W. is a member of the advisory board of HiberCell. Other authors declare no conflicts.
Figures






Update of
-
Cap-independent translation directs stress-induced differentiation of the protozoan parasite Toxoplasma gondii.bioRxiv [Preprint]. 2024 Sep 18:2024.09.17.613578. doi: 10.1101/2024.09.17.613578. bioRxiv. 2024. Update in: J Biol Chem. 2024 Dec;300(12):107979. doi: 10.1016/j.jbc.2024.107979. PMID: 39345546 Free PMC article. Updated. Preprint.
References
-
- Castro Machado F., Bittencourt-Cunha P., Malvezzi A.M., Arico M., Radio S., Smircich P., et al. EIF2alpha phosphorylation is regulated in intracellular amastigotes for the generation of infective Trypanosoma cruzi trypomastigote forms. Cell. Microbiol. 2020;22 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
