

Mycobacterium tuberculosis Mce3R TetR-like Repressor Forms an Asymmetric Four-Helix Bundle and Binds a Nonpalindrome Sequence†
- PMID: 39545866
- PMCID: PMC11667970
- DOI: 10.1021/acschembio.4c00687
Mycobacterium tuberculosis Mce3R TetR-like Repressor Forms an Asymmetric Four-Helix Bundle and Binds a Nonpalindrome Sequence†
Abstract
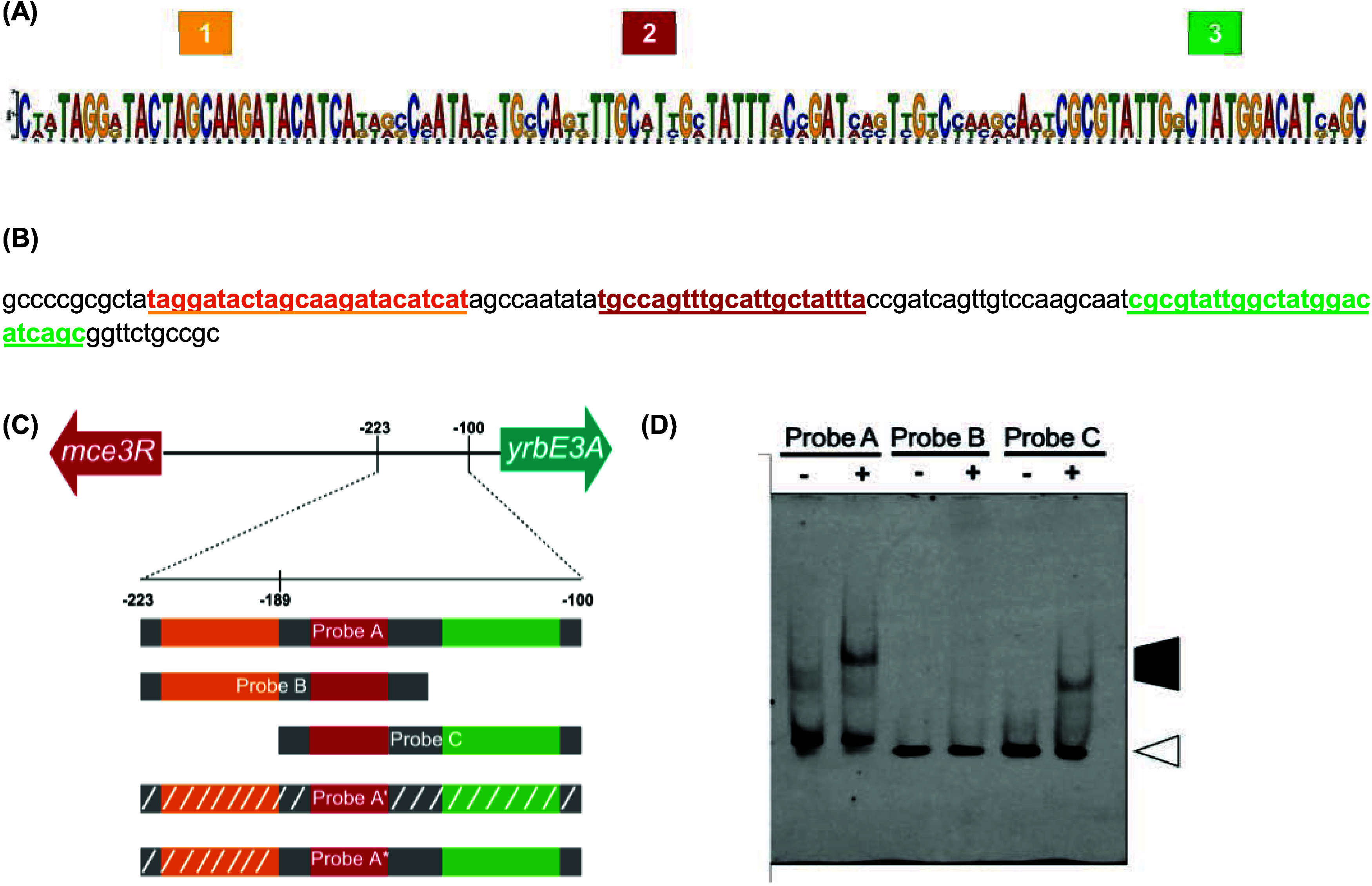
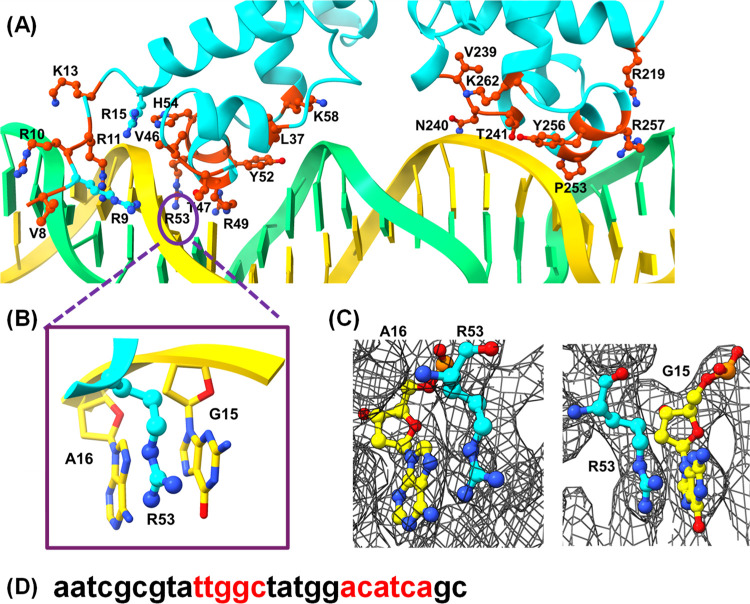
Mycobacterium tuberculosis (Mtb), the causative agent of tuberculosis, is a major global health concern. TetR family repressors (TFRs) are important for Mtb's adaptation to the human host environment. Our study focuses on one notable Mtb repressor, Mce3R, composed of an unusual double TFR motif. Mce3R-regulated genes encode enzymes implicated in cholesterol metabolism, resistance against reactive oxygen species, and lipid transport activities important for Mtb survival and persistence in the host and for the cellular activity of a 6-azasteroid derivative. Here, we present the structure of Mce3R bound to its DNA operator, unveiling a unique asymmetric assembly previously unreported. We obtained a candidate DNA-binding motif through MEME motif analysis, comparing intergenic regions of mce3R orthologues and identifying nonpalindromic regions conserved between orthologues. Using an electrophoretic mobility shift assay (EMSA), we confirmed that Mce3R binds to a 123-bp sequence that includes the predicted motif. Using scrambled DNA and DNA oligonucleotides of varying lengths with sequences from the upstream region of the yrbE3A (mce3) operon, we elucidated the operator region to be composed of two Mce3R binding sites, each a 25-bp asymmetric sequence separated by 53 bp. Mce3R binds with a higher affinity to the downstream site with a Kd of 2.4 ± 0.7 nM. The cryo-EM structure of Mce3R bound to the 123-bp sequence was refined to a resolution of 2.51 Å. Each Mce3R monomer comprises 21 α-helices (α1-α21) folded into an asymmetric TFR-like structure with a core asymmetric four-helix bundle. This complex has two nonidentical HTH motifs and a single ligand-binding domain. The two nonidentical HTHs from each TFR bind within the high-affinity, nonpalindromic operator motif, with Arg53 and Lys262 inserted into the major groove. Site-directed mutagenesis of Arg53 to alanine abrogated DNA binding, validating the Mce3R/DNA structure obtained. Among 811,645 particles, 63% were Mce3R homodimer bound to two duplex oligonucleotides. Mce3R homodimerizes primarily through α15, and each monomer binds to an identical site in the DNA duplex oligonucleotide.
Conflict of interest statement
The authors declare no competing financial interest.
Figures








Similar articles
-
Mce3R, a TetR-type transcriptional repressor, controls the expression of a regulon involved in lipid metabolism in Mycobacterium tuberculosis.Microbiology (Reading). 2009 Jul;155(Pt 7):2245-2255. doi: 10.1099/mic.0.027086-0. Epub 2009 Apr 23. Microbiology (Reading). 2009. PMID: 19389781
-
Structural and functional characterization of a ketosteroid transcriptional regulator of Mycobacterium tuberculosis.J Biol Chem. 2015 Jan 9;290(2):872-82. doi: 10.1074/jbc.M114.607481. Epub 2014 Nov 18. J Biol Chem. 2015. PMID: 25406313 Free PMC article.
-
The structure of the arginine repressor from Mycobacterium tuberculosis bound with its DNA operator and Co-repressor, L-arginine.J Mol Biol. 2009 Apr 24;388(1):85-97. doi: 10.1016/j.jmb.2009.02.053. Epub 2009 Mar 2. J Mol Biol. 2009. PMID: 19265706
-
Study of the role of Mce3R on the transcription of mce genes of Mycobacterium tuberculosis.BMC Microbiol. 2008 Feb 27;8:38. doi: 10.1186/1471-2180-8-38. BMC Microbiol. 2008. PMID: 18304349 Free PMC article.
-
The TetR family of transcriptional repressors.Microbiol Mol Biol Rev. 2005 Jun;69(2):326-56. doi: 10.1128/MMBR.69.2.326-356.2005. Microbiol Mol Biol Rev. 2005. PMID: 15944459 Free PMC article. Review.
References
-
- WHO . Global Tuberculosis Report 2023 2023www.who.inthttps://www.who.int/teams/global-tuberculosis-programme/tb-re.... (accessed July 4, 2024).
-
- Anand S.; Singh V.; Singh A. K.; Mittal M.; Datt M.; Subramani B.; S K. Equilibrium binding and kinetic characterization of putative tetracycline repressor family transcription regulator Fad35R from Mycobacterium tuberculosis. FEBS J. 2012, 279 (17), 3214–3228. 10.1111/j.1742-4658.2012.08707.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
