

Cell response to extracellular matrix viscous energy dissipation outweighs high-rigidity sensing
- PMID: 39546608
- PMCID: PMC11567001
- DOI: 10.1126/sciadv.adf9758
Cell response to extracellular matrix viscous energy dissipation outweighs high-rigidity sensing
Abstract
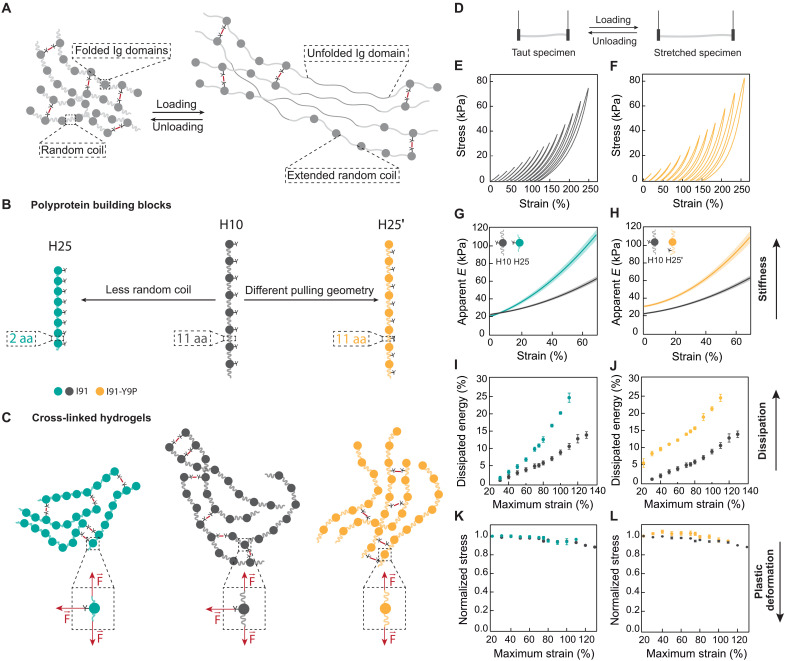
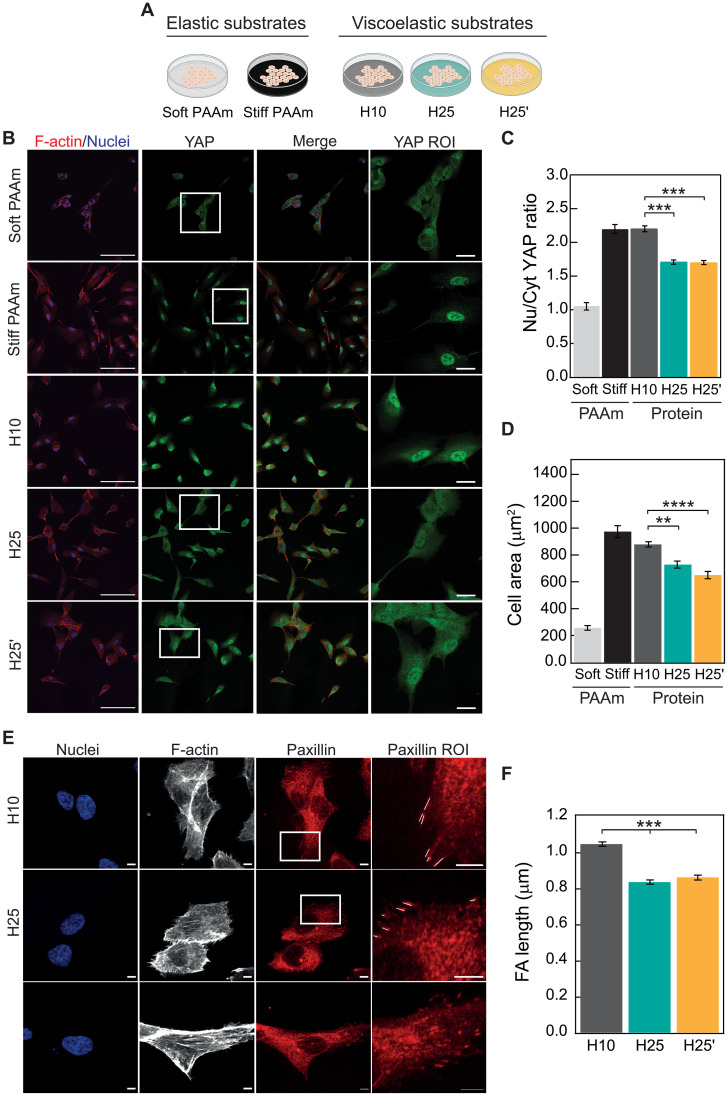
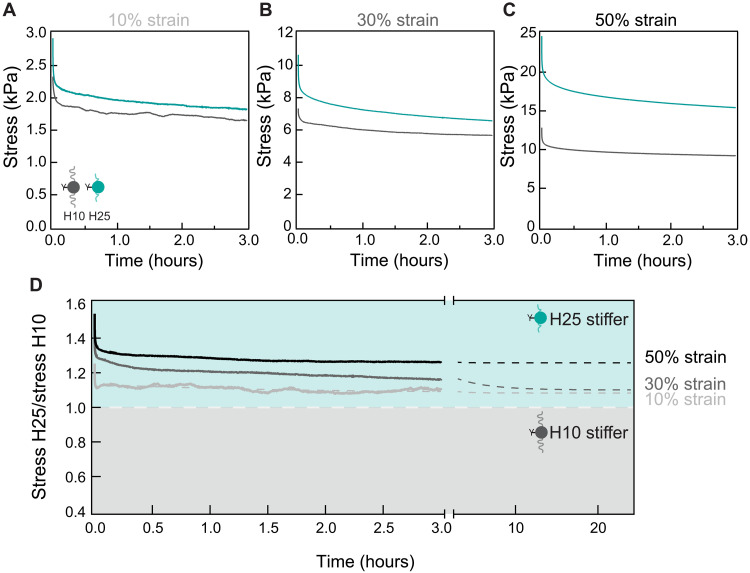
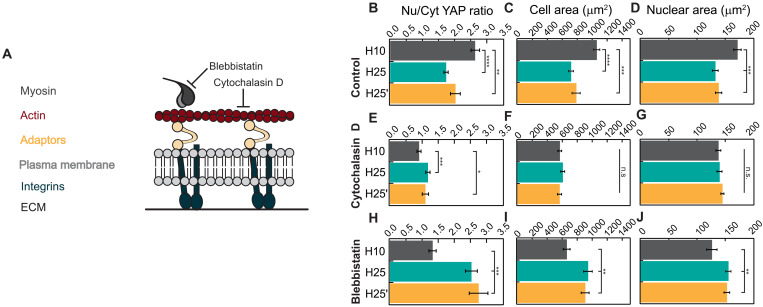
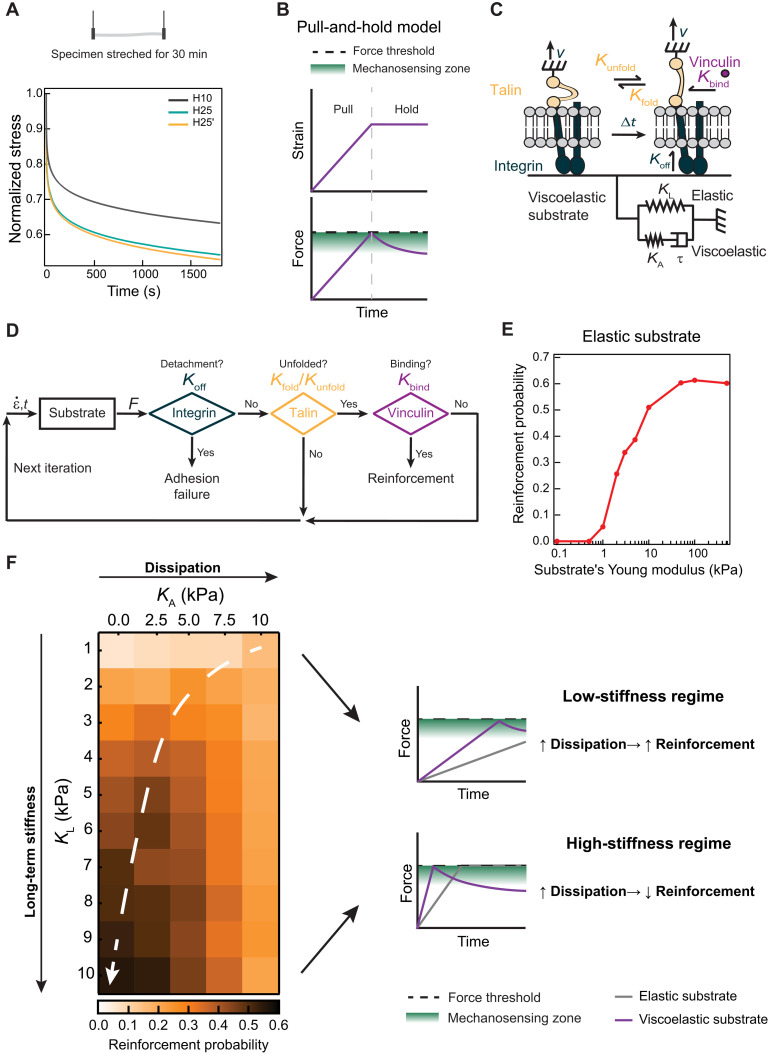
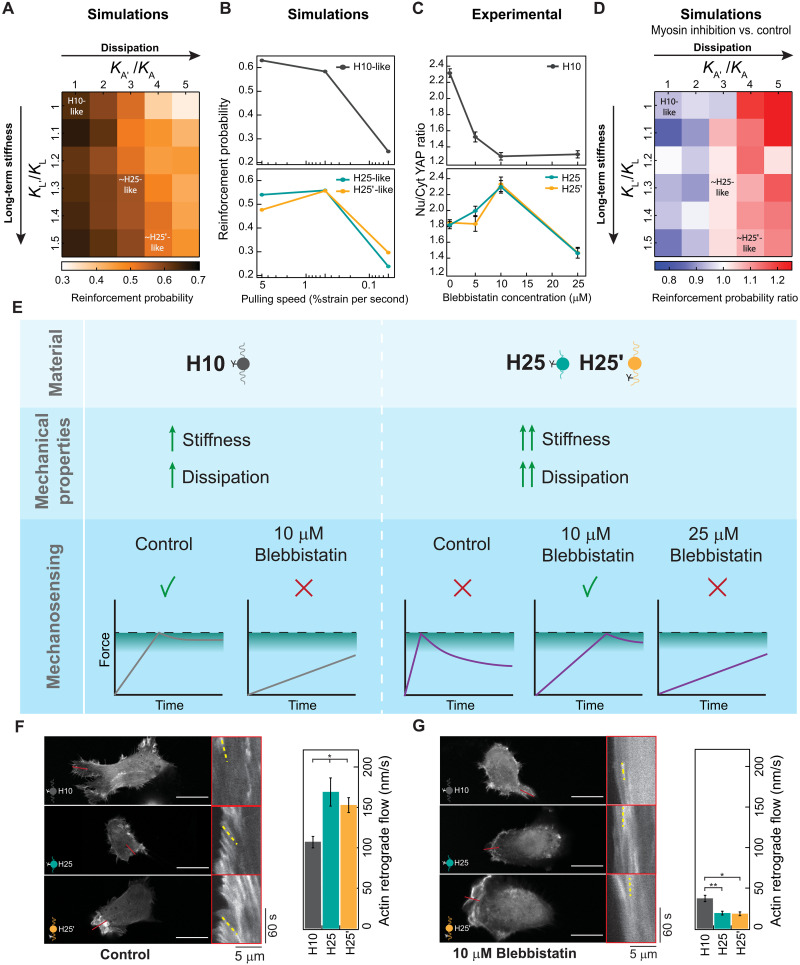
The mechanics of the extracellular matrix (ECM) determine cell activity and fate through mechanoresponsive proteins including Yes-associated protein 1 (YAP). Rigidity and viscous relaxation have emerged as the main mechanical properties of the ECM steering cell behavior. However, how cells integrate coexisting ECM rigidity and viscosity cues remains poorly understood, particularly in the high-stiffness regime. Here, we have exploited engineered stiff viscoelastic protein hydrogels to show that, contrary to current models of cell-ECM interaction, substrate viscous energy dissipation attenuates mechanosensing even when cells are exposed to higher effective rigidity. This unexpected behavior is however readily captured by a pull-and-hold model of molecular clutch-based cell mechanosensing, which also recapitulates opposite cellular response at low rigidities. Consistent with predictions of the pull-and-hold model, we find that myosin inhibition can boost mechanosensing on cells cultured on dissipative matrices. Together, our work provides general mechanistic understanding on how cells respond to the viscoelastic properties of the ECM.
Figures
References
-
- Engler A. J., Sen S., Sweeney H. L., Discher D. E., Matrix elasticity directs stem cell lineage specification. Cell 126, 677–689 (2006). - PubMed
-
- Elosegui-Artola A., Trepat X., Roca-Cusachs P., Control of mechanotransduction by molecular clutch dynamics. Trends Cell Biol. 28, 356–367 (2018). - PubMed
-
- Segel M., Neumann B., Hill M. F. E., Weber I. P., Viscomi C., Zhao C., Young A., Agley C. C., Thompson A. J., Gonzalez G. A., Sharma A., Holmqvist S., Rowitch D. H., Franze K., Franklin R. J. M., Chalut K. J., Niche stiffness underlies the ageing of central nervous system progenitor cells. Nature 573, 130–134 (2019). - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
