

PKA regulates autophagy through lipolysis during fasting
- PMID: 39547583
- PMCID: PMC11697058
- DOI: 10.1016/j.mocell.2024.100149
PKA regulates autophagy through lipolysis during fasting
Abstract
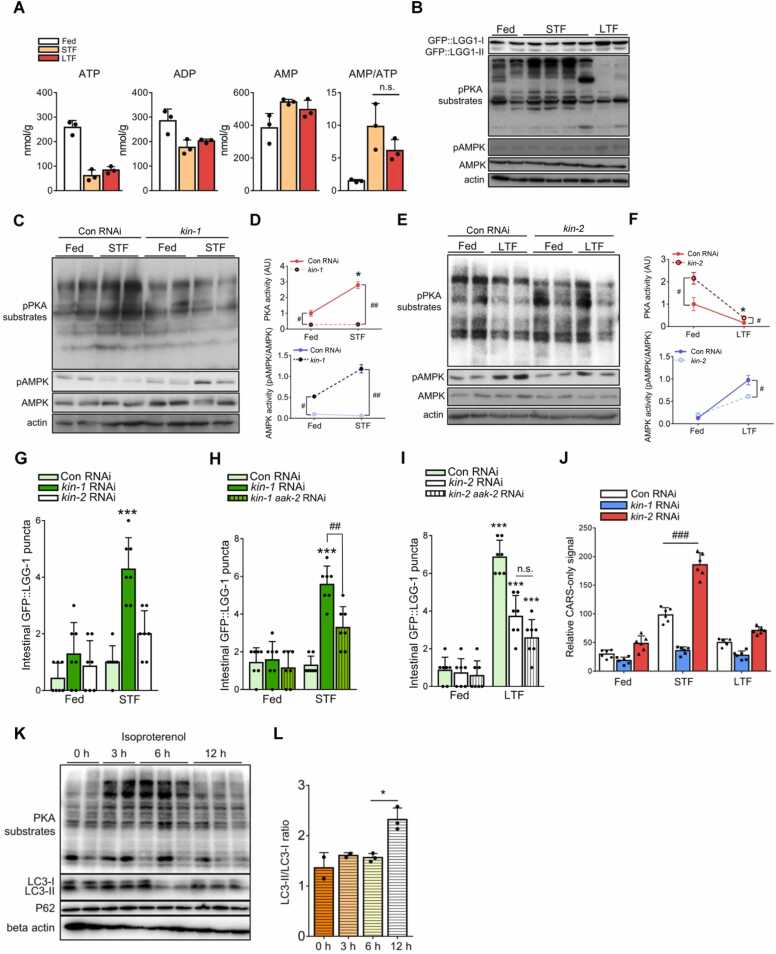
Autophagy is a crucial intracellular degradation process that provides energy and supports nutrient deprivation adaptation. However, the mechanisms by which these cells detect lipid scarcity and regulate autophagy are poorly understood. In this study, we demonstrate that protein kinase A (PKA)-dependent lipolysis delays autophagy initiation during short-term nutrient deprivation by inhibiting AMP-activated protein kinase (AMPK). Using coherent anti-Stokes Raman spectroscopy, we visualized free fatty acids (FFAs) in vivo and observed that lipolysis-derived FFAs were used before the onset of autophagy. Our data suggest that autophagy is triggered when the supply of FFAs is insufficient to meet energy demands. Furthermore, PKA activation promotes lipolysis and suppresses AMPK-driven autophagy during early fasting. Disruption of this regulatory axis impairs motility and reduces the lifespan of Caenorhabditis elegans during fasting. These findings establish PKA as a critical regulator of catabolic pathways, prioritizing lipolysis over autophagy by modulating AMPK activity to prevent premature autophagic degradation during transient nutrient deprivation.
Keywords: AMP-activated protein kinase; Autophagy; Caenorhabditis elegans; Free fatty acid; Lipolysis; Protein kinase A.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interests The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
