

Loss of cytoplasmic actin filaments raises nuclear actin levels to drive INO80C-dependent chromosome fragmentation
- PMID: 39548059
- PMCID: PMC11568269
- DOI: 10.1038/s41467-024-54141-0
Loss of cytoplasmic actin filaments raises nuclear actin levels to drive INO80C-dependent chromosome fragmentation
Abstract
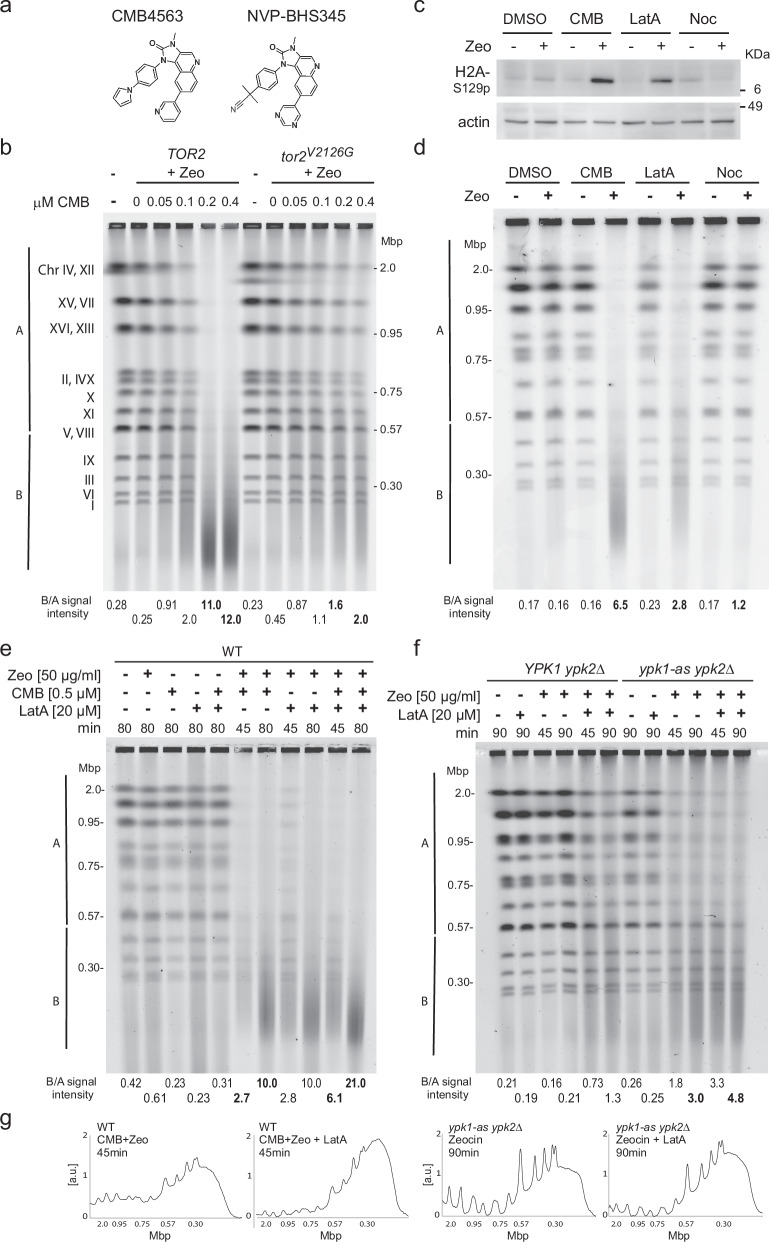
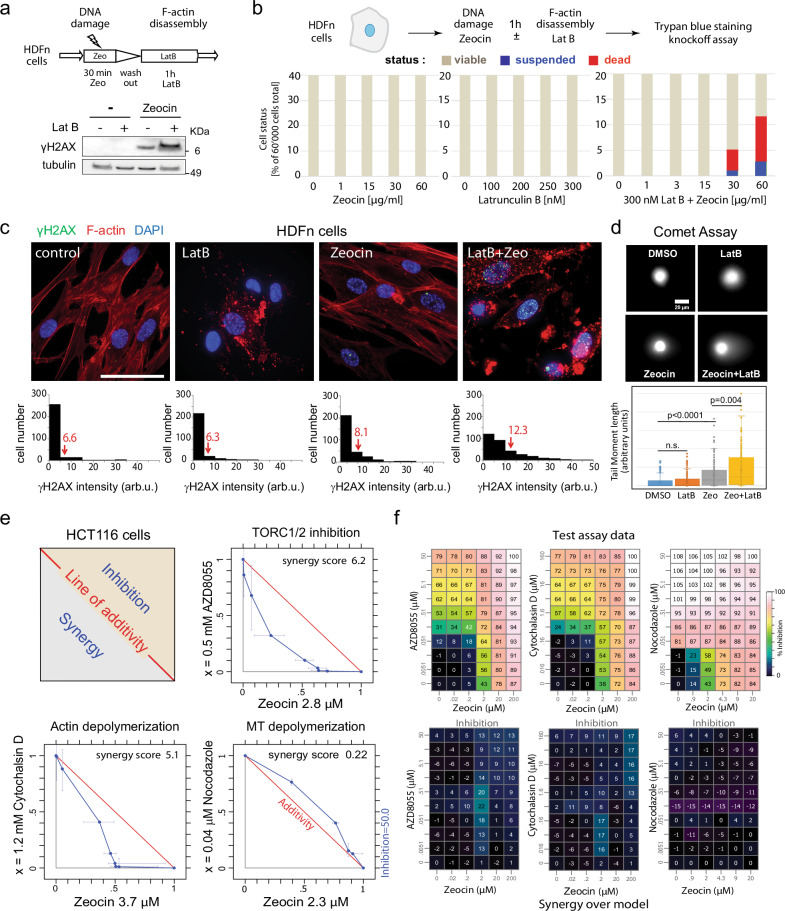
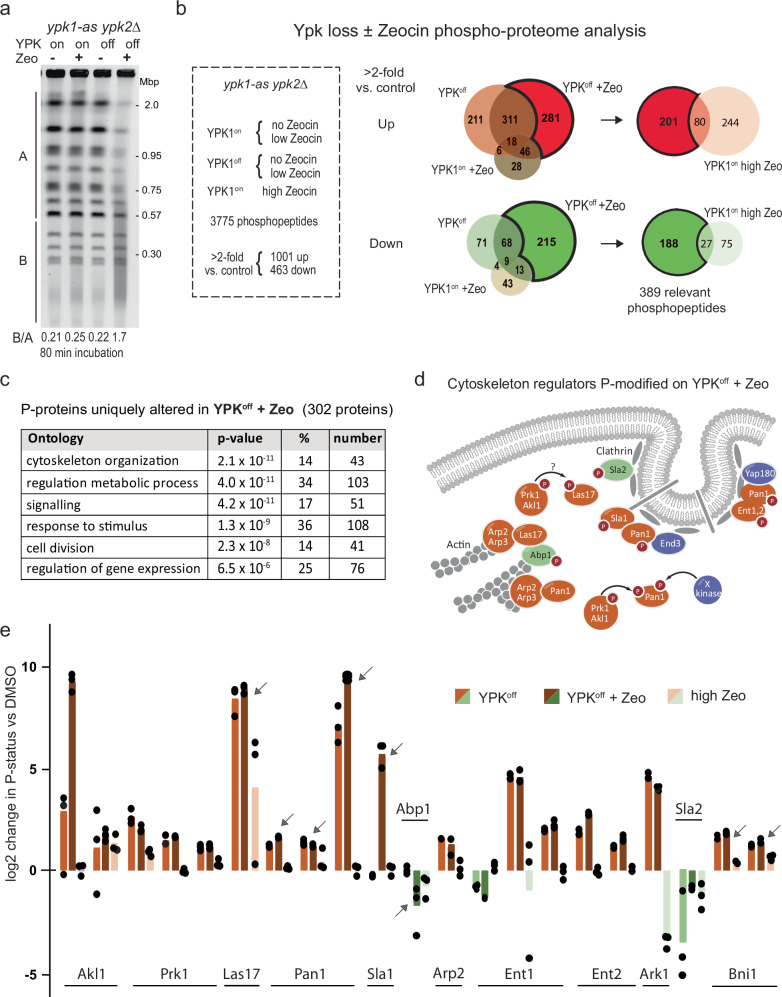
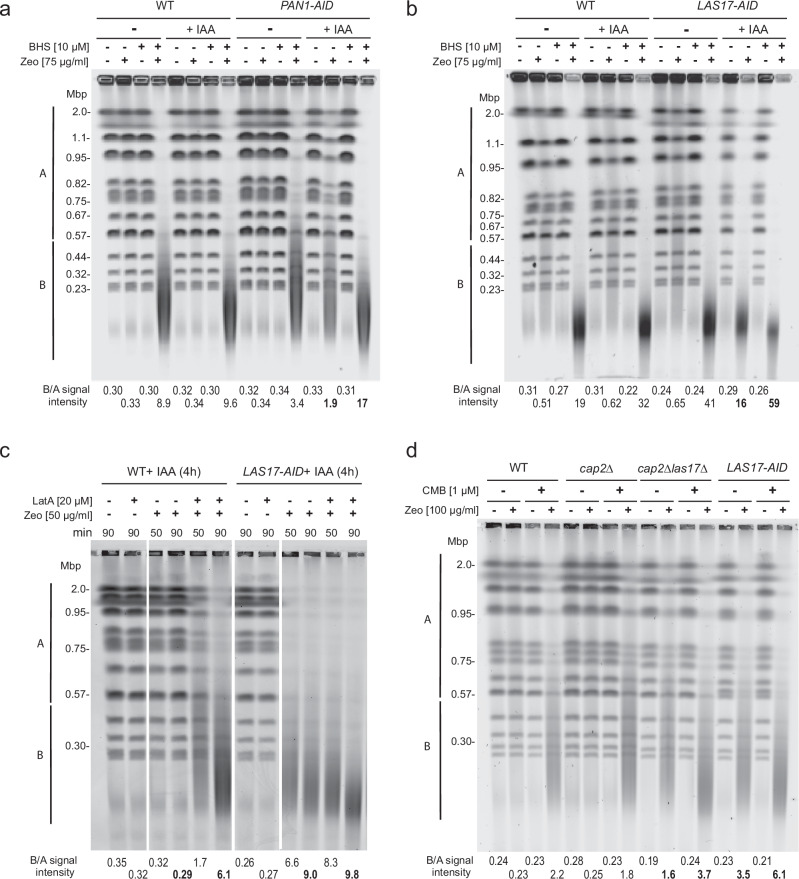
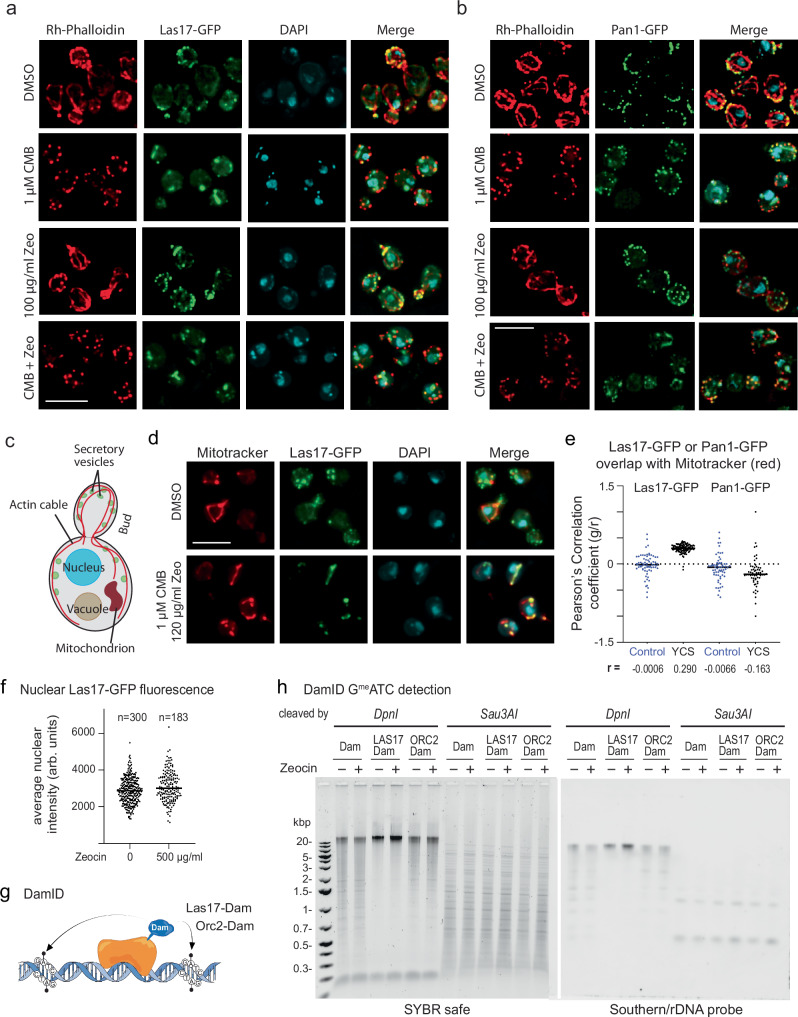
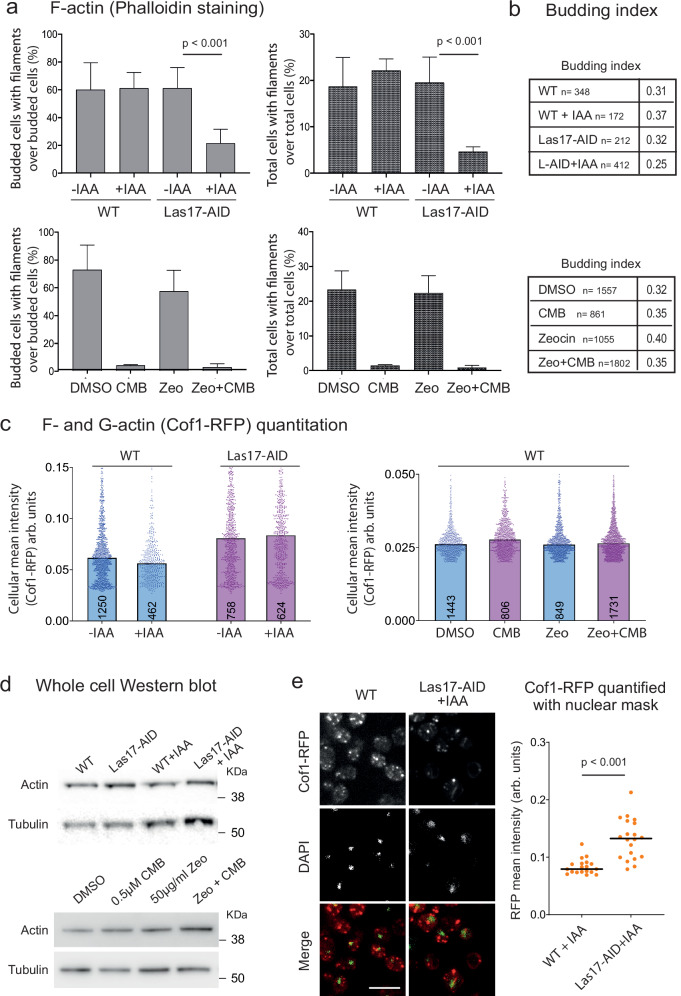
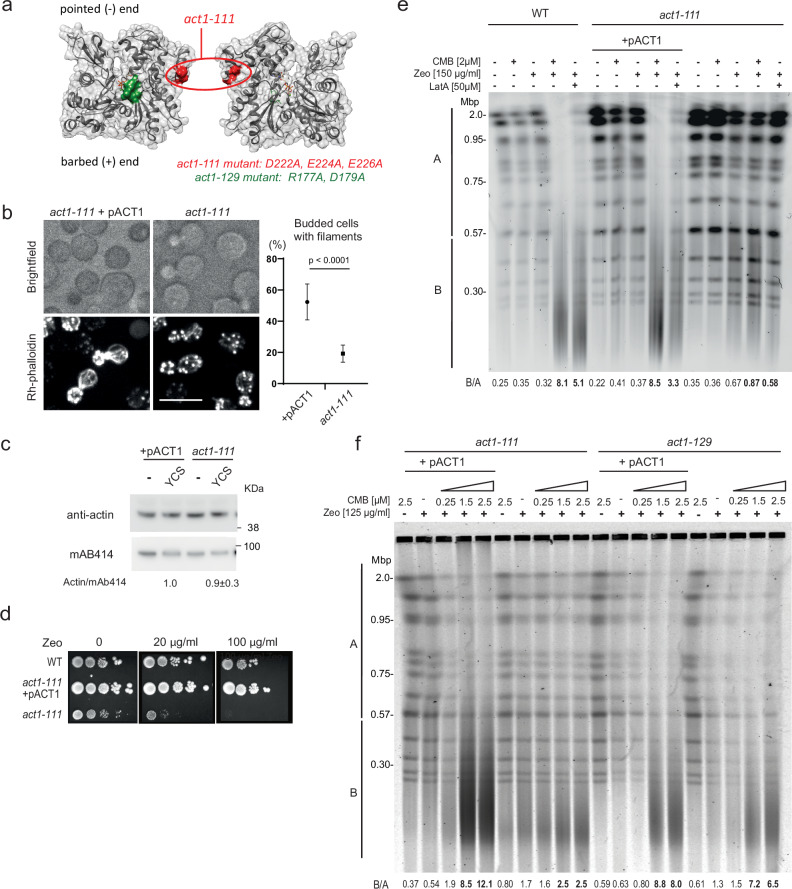
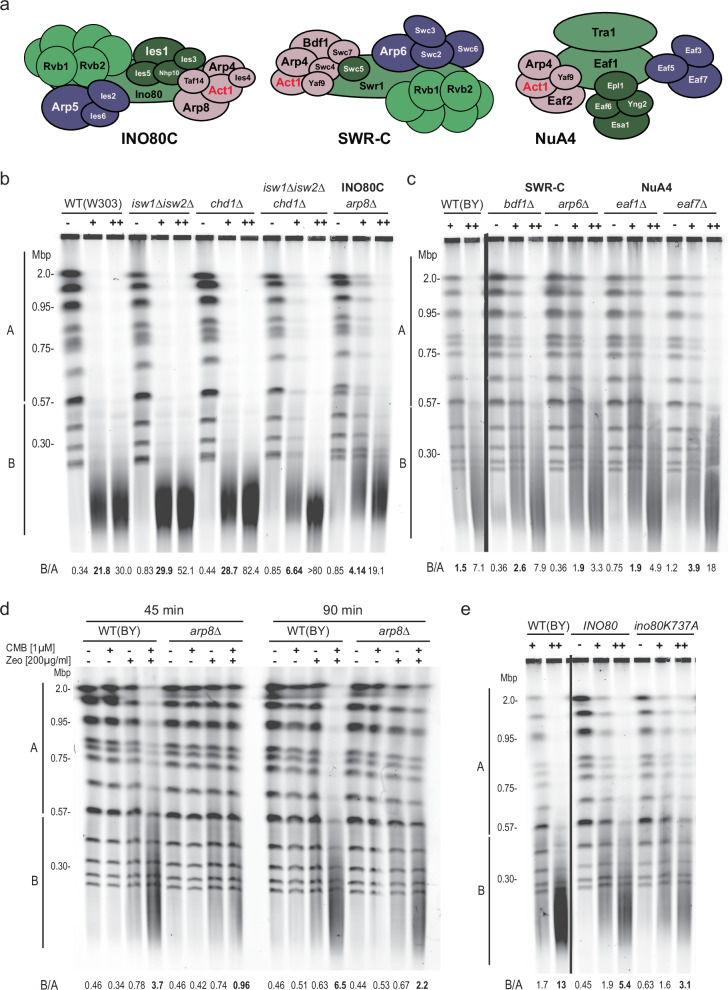
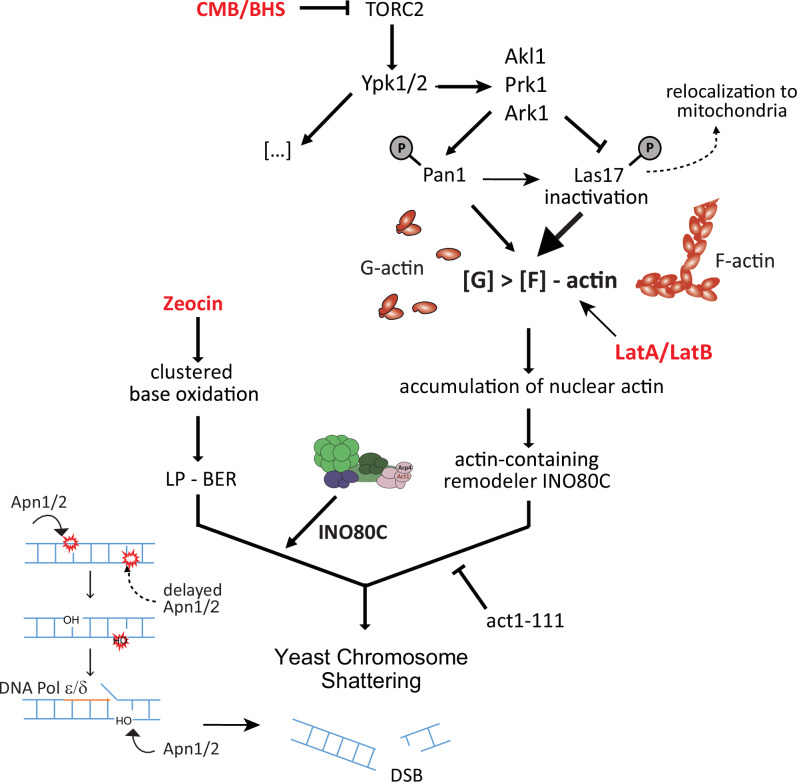
Loss of cytosolic actin filaments upon TORC2 inhibition triggers chromosome fragmentation in yeast, which results from altered base excision repair of Zeocin-induced lesions. To find the link between TORC2 kinase and this yeast chromosome shattering (YCS) we performed phosphoproteomics. YCS-relevant phospho-targets included plasma membrane-associated regulators of actin polymerization, such as Las17, the yeast Wiscott-Aldrich Syndrome protein. Induced degradation of Las17 was sufficient to trigger YCS in presence of Zeocin, bypassing TORC2 inhibition. In yeast, Las17 does not act directly at damage, but instead its loss, like TORC2 inhibition, raises nuclear actin levels. Nuclear actin, in complex with Arp4, forms an essential subunit of several nucleosome remodeler complexes, including INO80C, which facilitates DNA polymerase elongation. Here we show that the genetic ablation of INO80C activity leads to partial YCS resistance, suggesting that elevated levels of nuclear G-actin may stimulate INO80C to increase DNA polymerase processivity and convert single-strand lesions into double-strand breaks.
© 2024. The Author(s).
Conflict of interest statement
Figures
References
-
- Vartiainen, M. K., Guettler, S., Larijani, B. & Treisman, R. Nuclear actin regulates dynamic subcellular localization and activity of the SRF cofactor MAL. Science316, 1749–1752 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
