

TORC2 inhibition triggers yeast chromosome fragmentation through misregulated Base Excision Repair of clustered oxidation events
- PMID: 39548071
- PMCID: PMC11568337
- DOI: 10.1038/s41467-024-54142-z
TORC2 inhibition triggers yeast chromosome fragmentation through misregulated Base Excision Repair of clustered oxidation events
Abstract
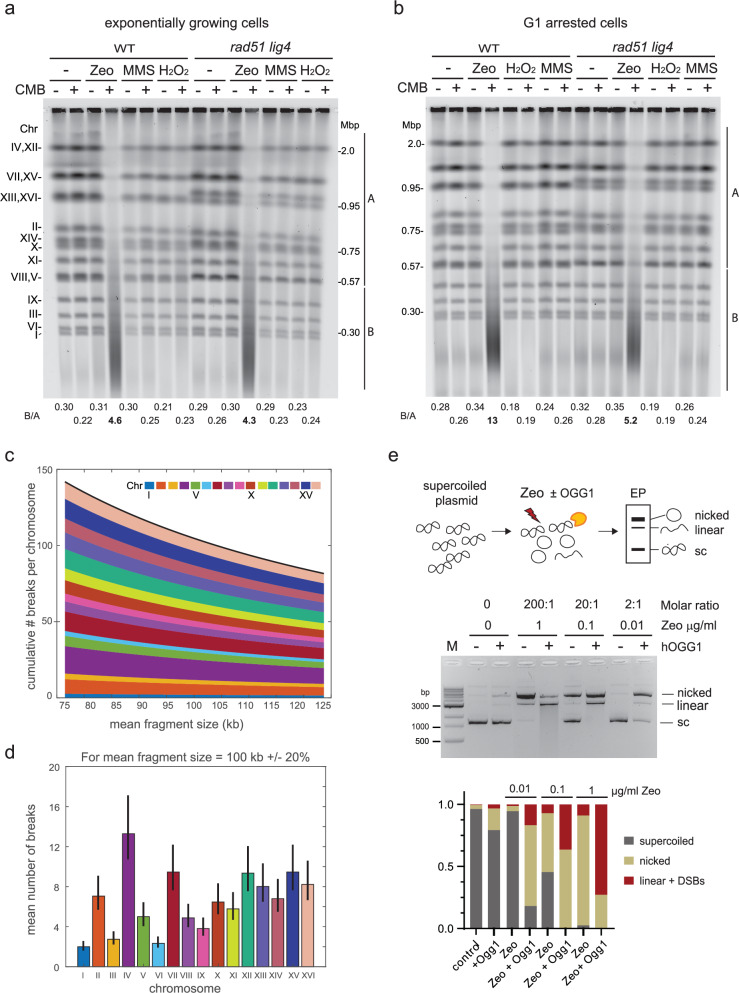
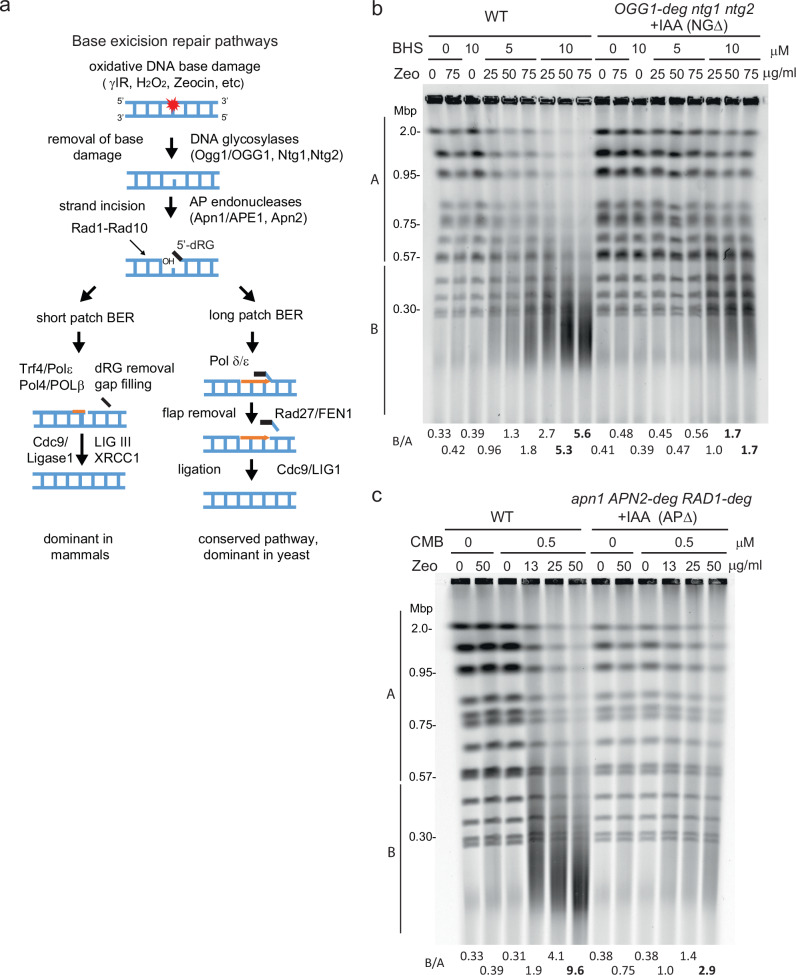
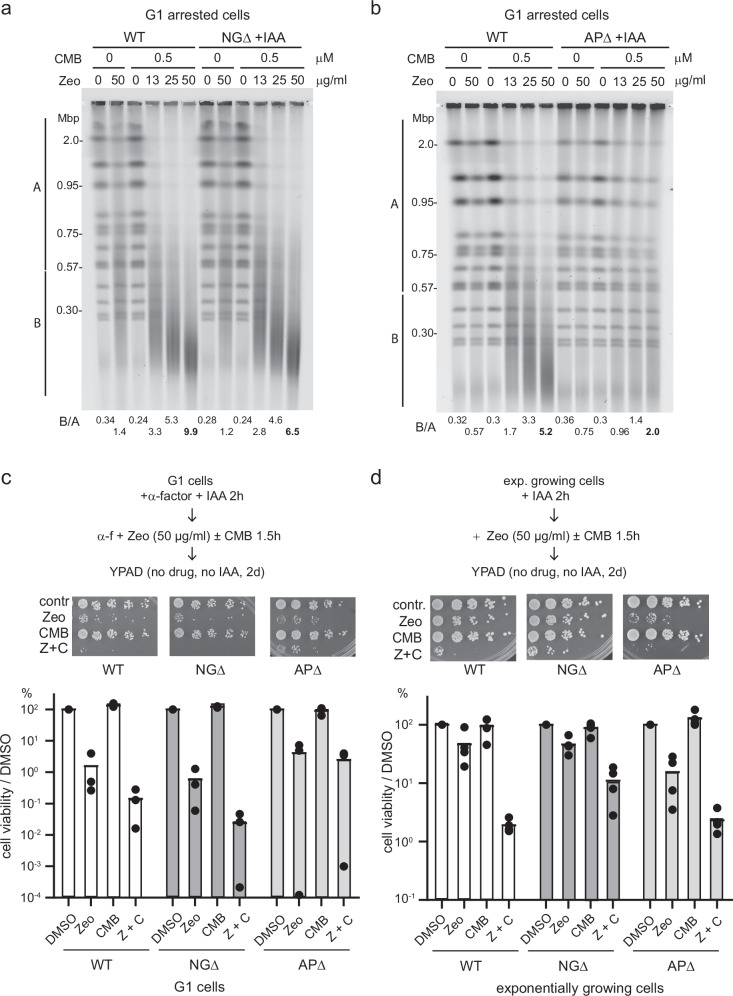
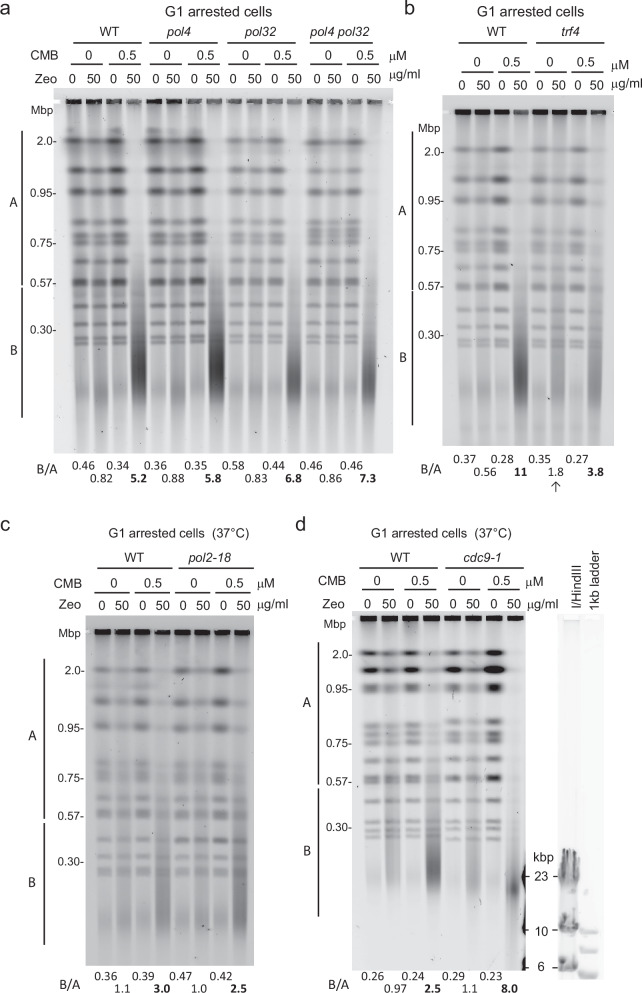
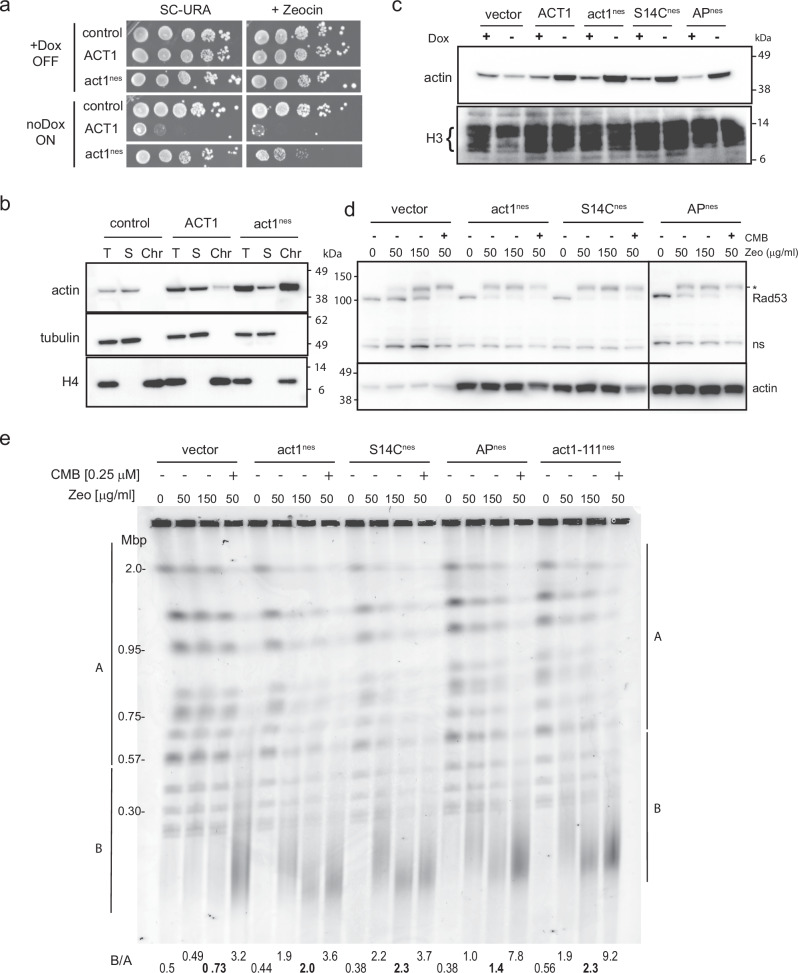
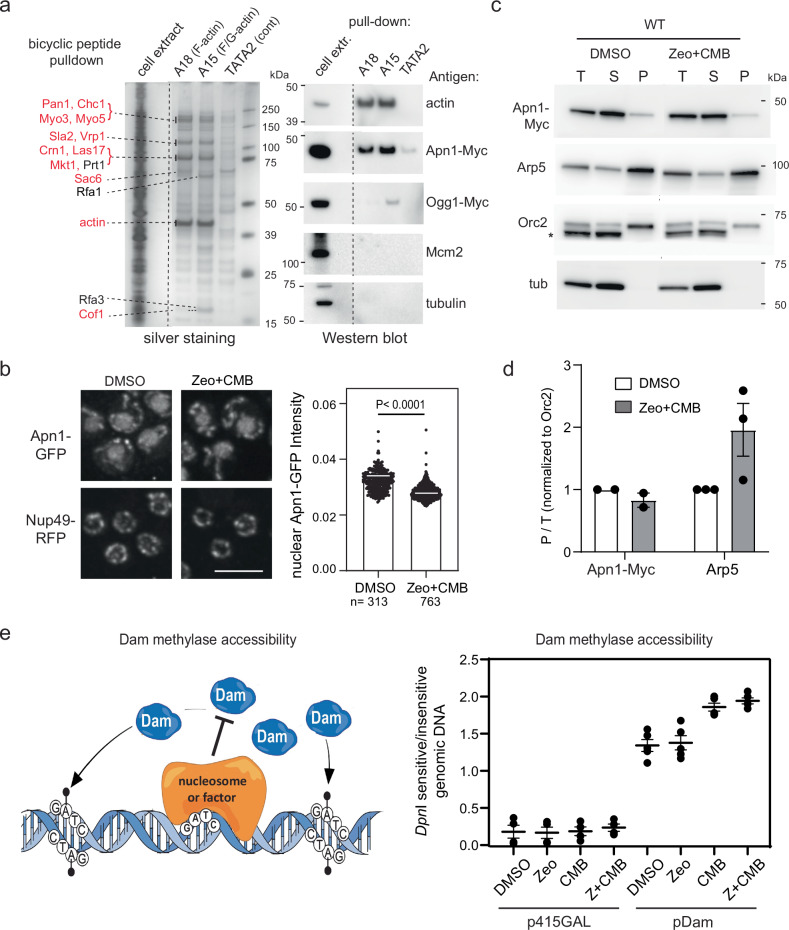
Combinational therapies provoking cell death are of major interest in oncology. Combining TORC2 kinase inhibition with the radiomimetic drug Zeocin results in a rapid accumulation of double-strand breaks (DSB) in the budding yeast genome. This lethal Yeast Chromosome Shattering (YCS) requires conserved enzymes of base excision repair. YCS can be attenuated by eliminating three N-glycosylases or endonucleases Apn1/Apn2 and Rad1, which act to convert oxidized bases into abasic sites and single-strand nicks. Adjacent lesions must be repaired in a step-wise fashion to avoid generating DSBs. Artificially increasing nuclear actin by destabilizing cytoplasmic actin filaments or by expressing a nuclear export-deficient actin interferes with this step-wise repair and generates DSBs, while mutants that impair DNA polymerase processivity reduce them. Repair factors that bind actin include Apn1, RFA and the actin-dependent chromatin remodeler INO80C. During YCS, increased INO80C activity could enhance both DNA polymerase processivity and repair factor access to convert clustered lesions into DSBs.
© 2024. The Author(s).
Conflict of interest statement
Figures


References
-
- Lindahl, T. & Barnes, D. E. Repair of endogenous DNA damage. Cold Spring Harb. Symp. Quant. Biol.65, 127–133 (2000). - PubMed
-
- Methot, S. P. & Di Noia, J. M. Molecular mechanisms of somatic hypermutation and class switch recombination. Adv. Immunol.133, 37–87 (2017). - PubMed
-
- van Loon, B., Markkanen, E. & Hubscher, U. Oxygen as a friend and enemy: How to combat the mutational potential of 8-oxo-guanine. DNA Repair9, 604–616 (2010). - PubMed
-
- de la Torre-Ruiz, M. A., Pujol, N. & Sundaran, V. Coping with oxidative stress. The yeast model. Curr. Drug Targets16, 2–12 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
