

Functionally overlapping intra- and extralysosomal pathways promote bis(monoacylglycero)phosphate synthesis in mammalian cells
- PMID: 39548099
- PMCID: PMC11568333
- DOI: 10.1038/s41467-024-54213-1
Functionally overlapping intra- and extralysosomal pathways promote bis(monoacylglycero)phosphate synthesis in mammalian cells
Abstract
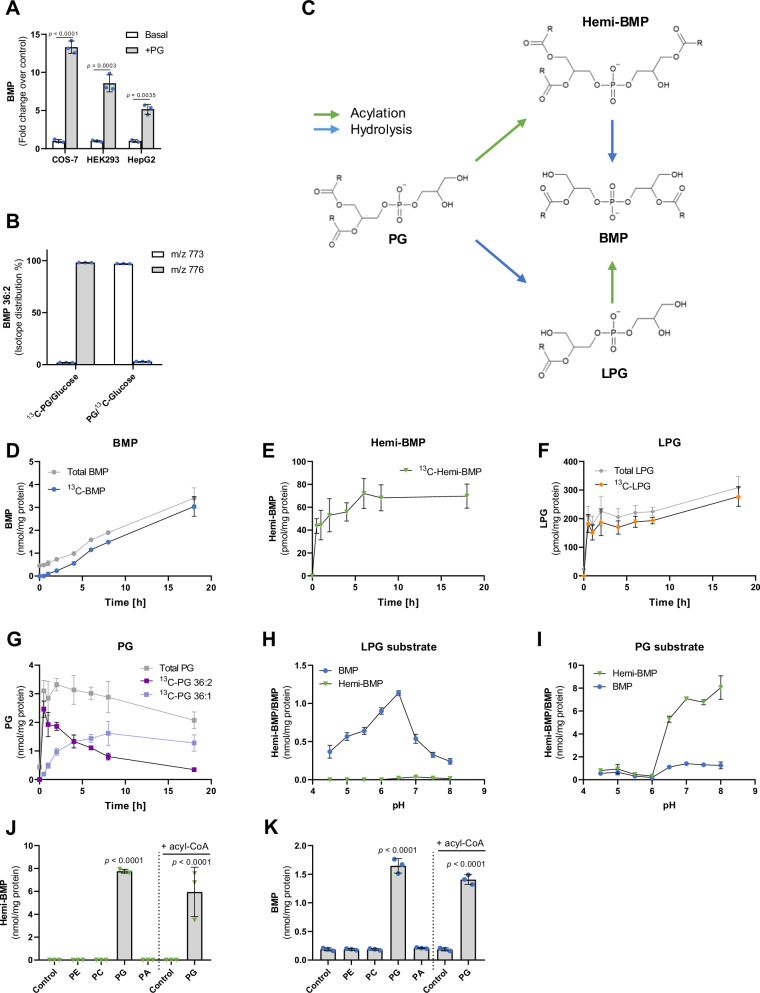
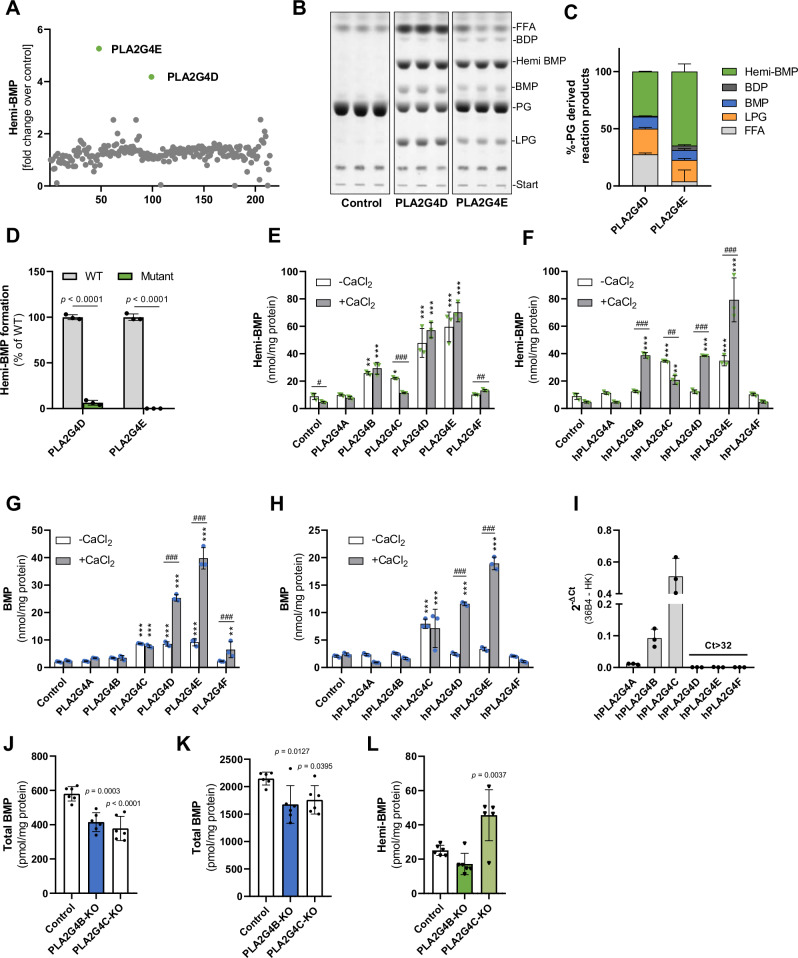
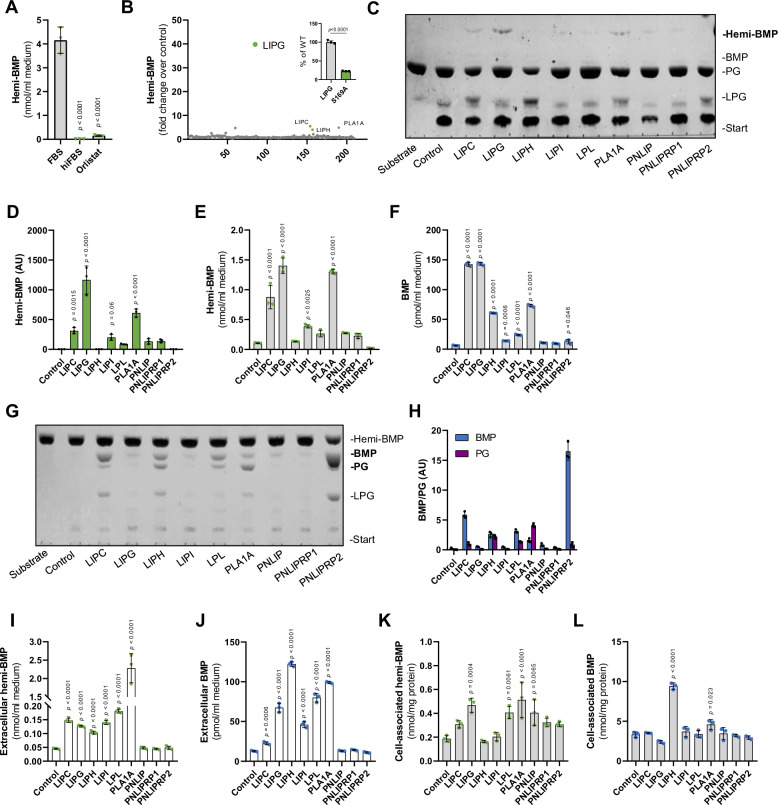
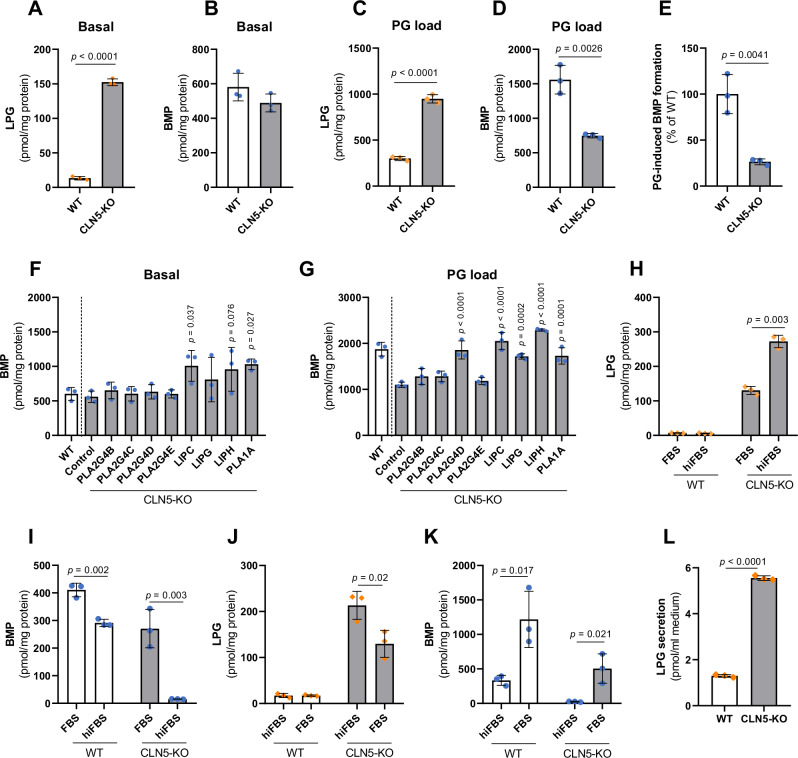
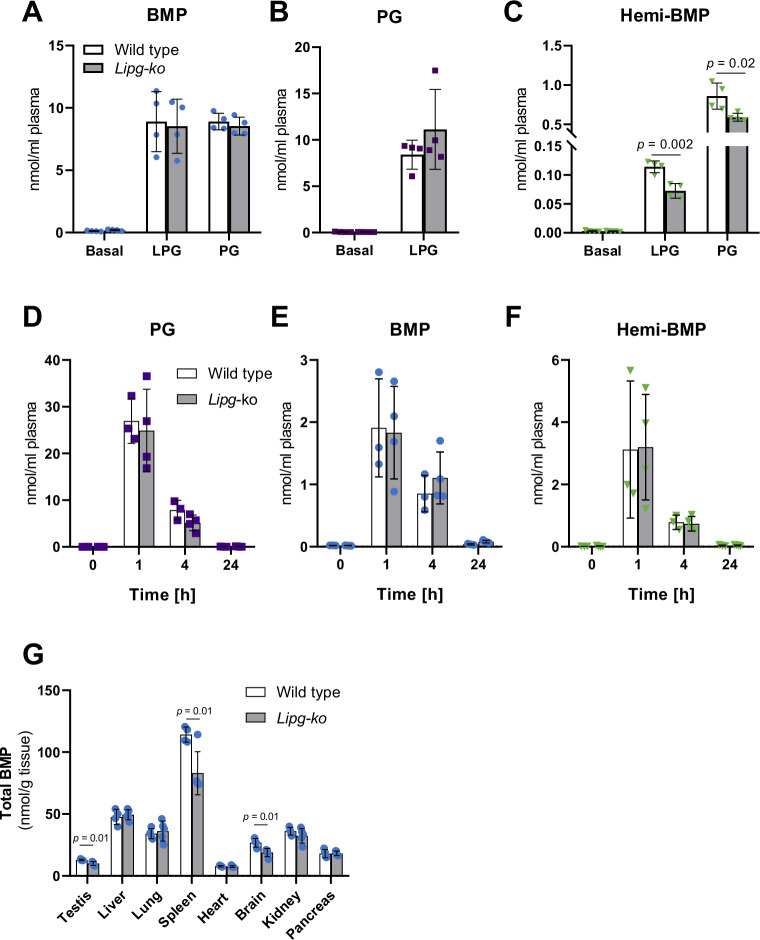
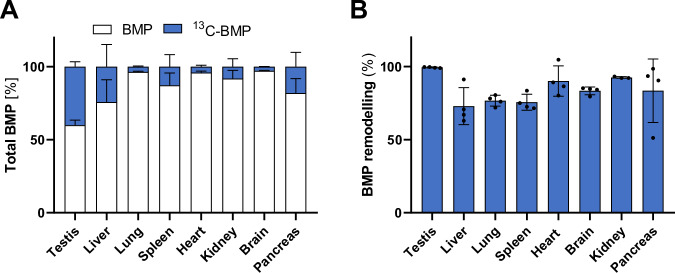
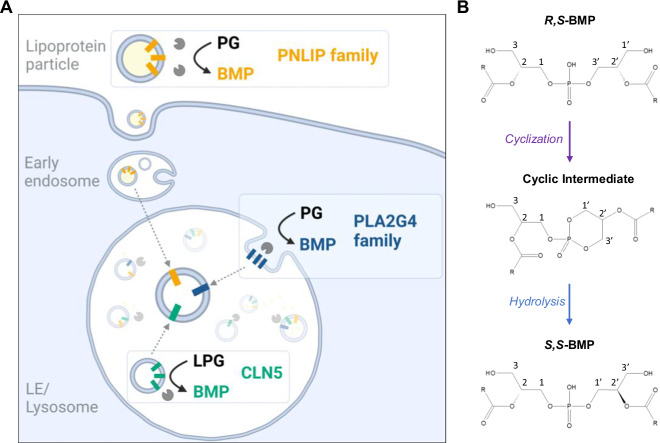
Bis(monoacylglycero)phosphate (BMP) is a major phospholipid constituent of intralumenal membranes in late endosomes/lysosomes, where it regulates the degradation and sorting of lipid cargo. Recent observations suggest that the Batten disease-associated protein CLN5 functions as lysosomal BMP synthase. Here, we show that transacylation reactions catalyzed by cytosolic and secreted enzymes enhance BMP synthesis independently of CLN5. The transacylases identified in this study are capable of acylating the precursor lipid phosphatidylglycerol (PG), generating acyl-PG, which is subsequently hydrolyzed to BMP. Extracellularly, acyl-PG and BMP are generated by endothelial lipase in cooperation with other serum enzymes of the pancreatic lipase family. The intracellular acylation of PG is catalyzed by several members of the cytosolic phospholipase A2 group IV (PLA2G4) family. Overexpression of secreted or cytosolic transacylases was sufficient to correct BMP deficiency in HEK293 cells lacking CLN5. Collectively, our observations suggest that functionally overlapping pathways promote BMP synthesis in mammalian cells.
© 2024. The Author(s).
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
