

Defective kinase activity of IKKα leads to combined immunodeficiency and disruption of immune tolerance in humans
- PMID: 39550372
- PMCID: PMC11569180
- DOI: 10.1038/s41467-024-54345-4
Defective kinase activity of IKKα leads to combined immunodeficiency and disruption of immune tolerance in humans
Abstract
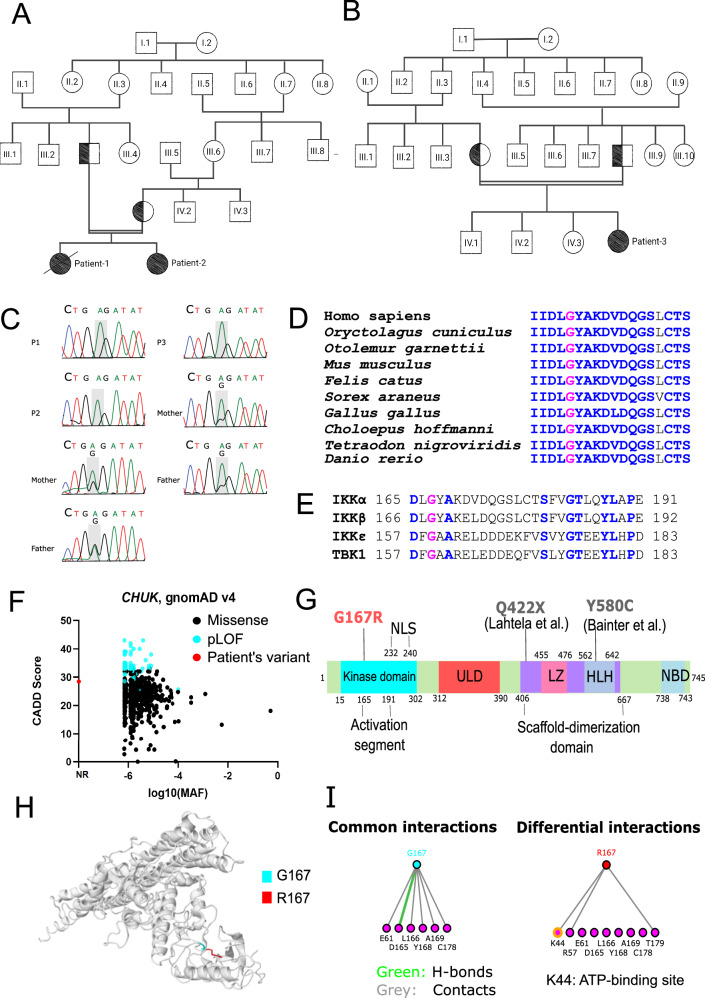
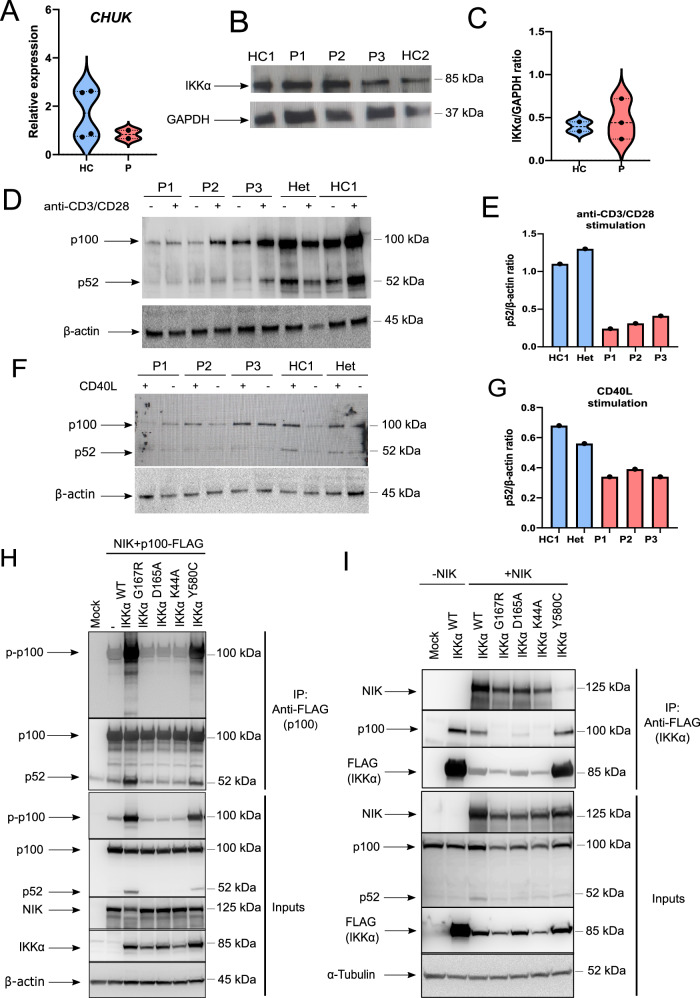
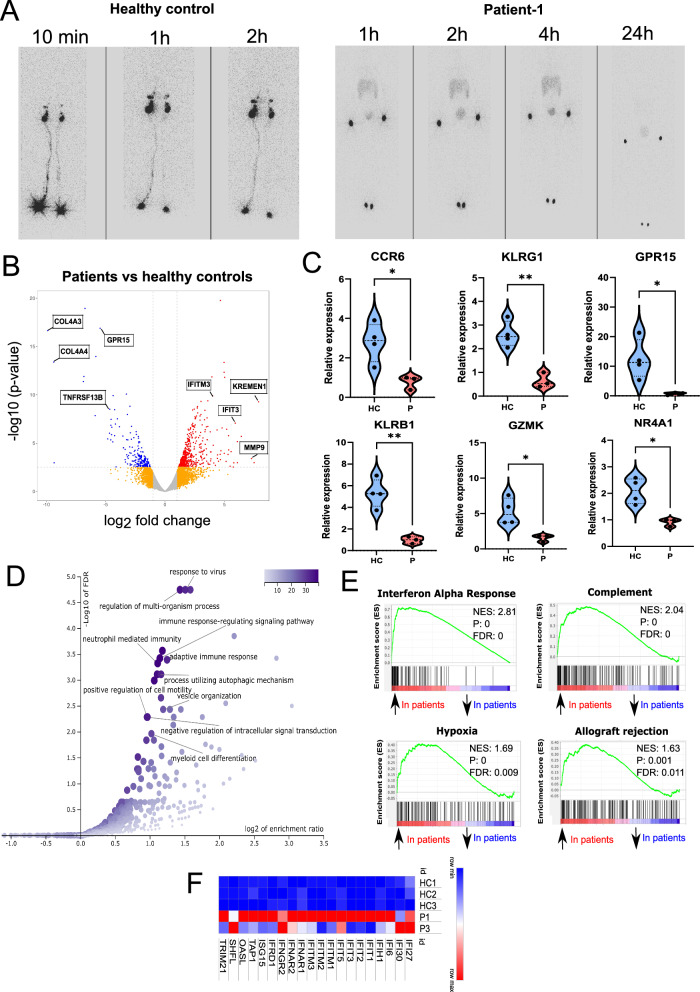
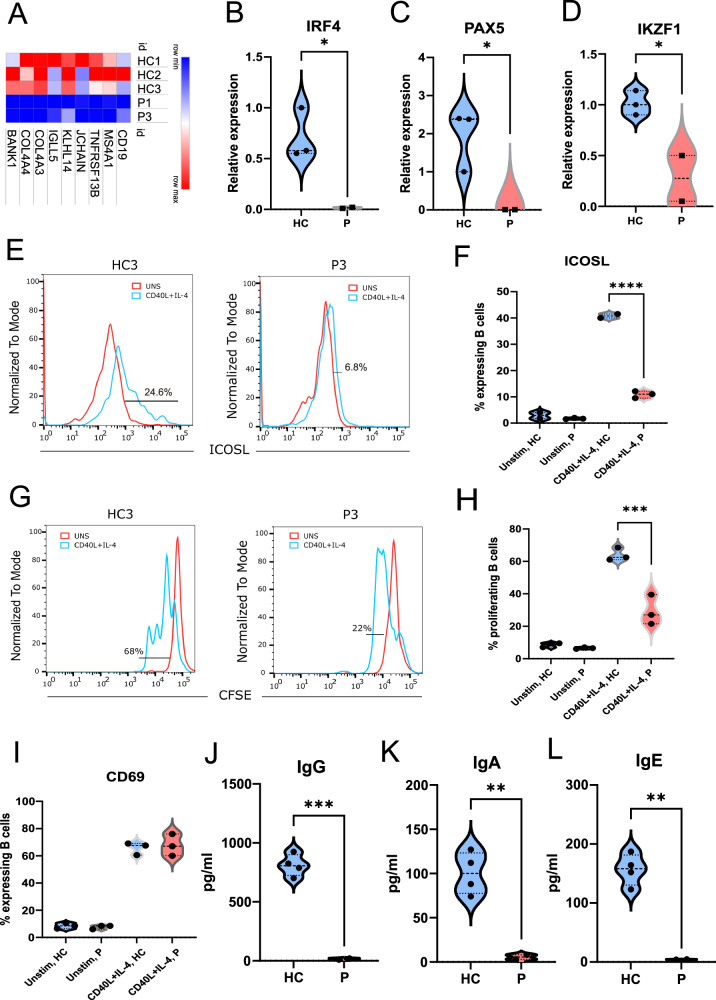
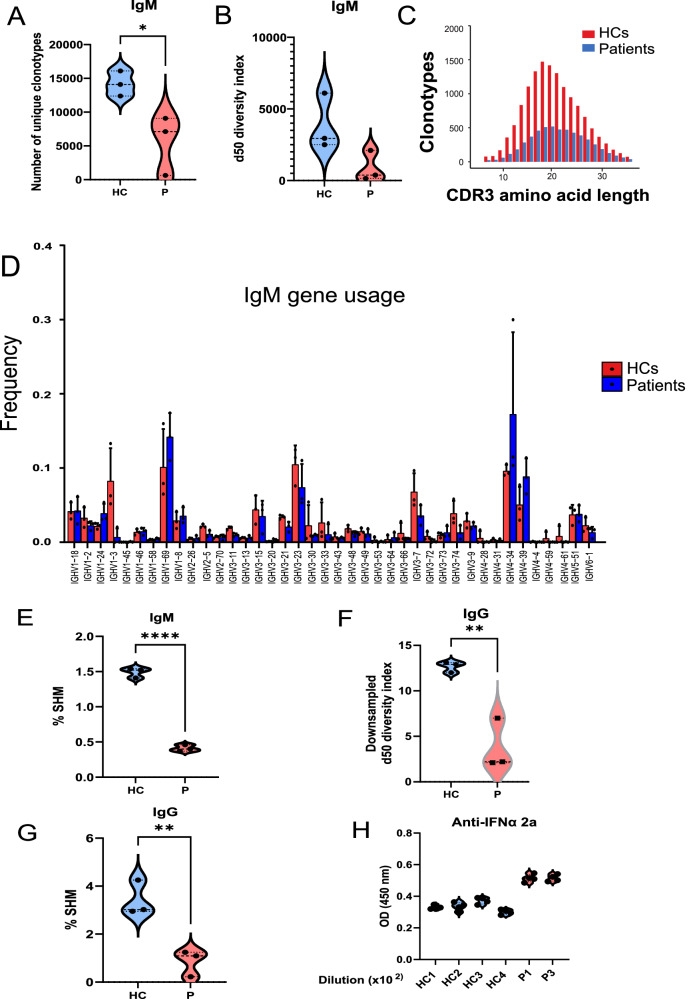
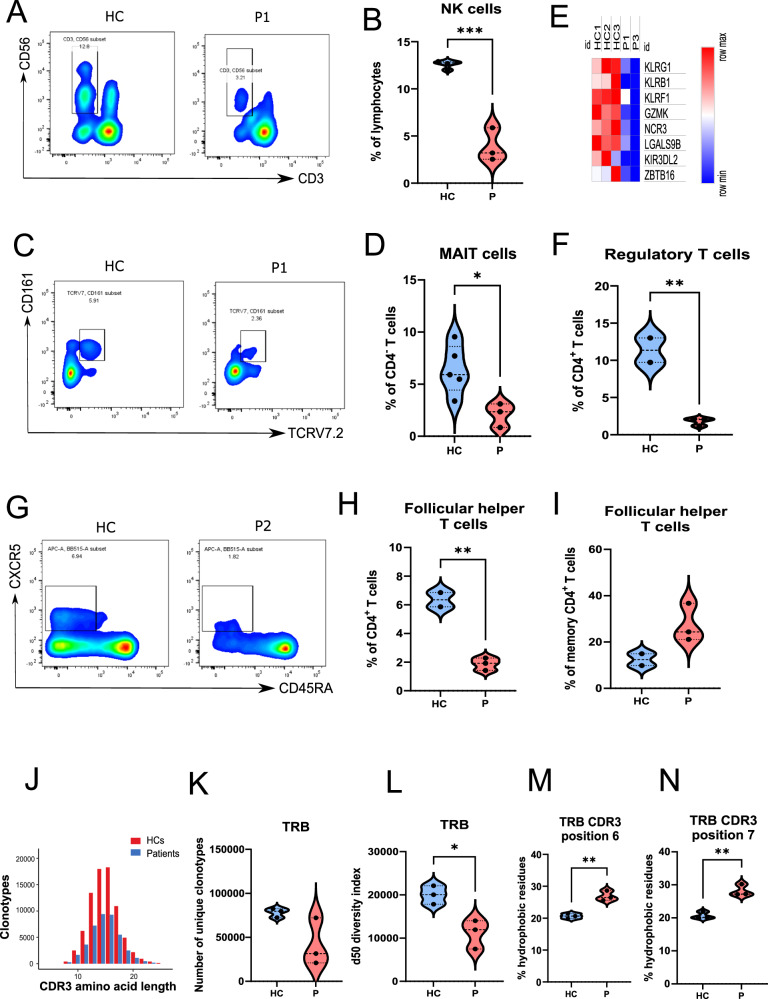
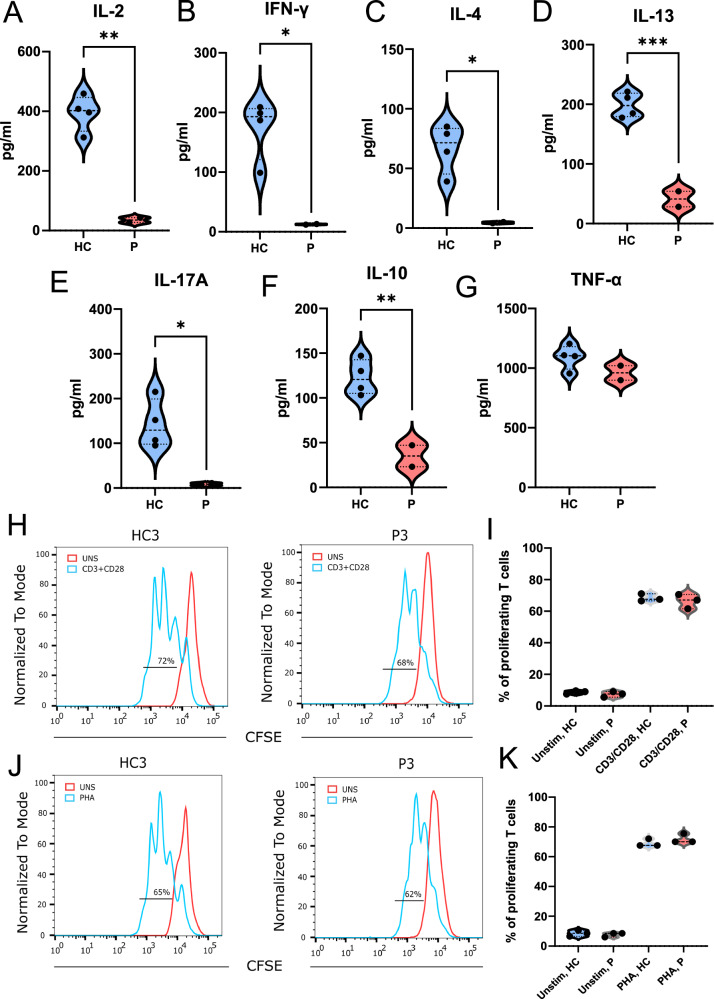
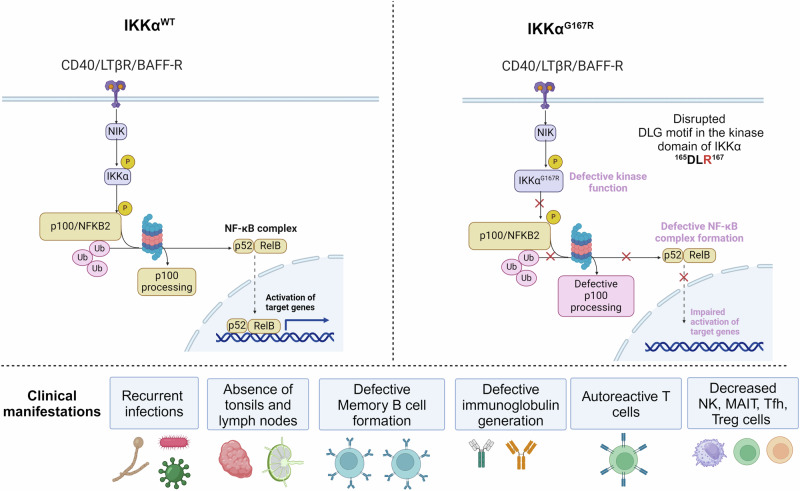
IKKα is a multifunctional serine/threonine kinase that controls various biological processes, either dependent on or independent of its kinase activity. However, the importance of the kinase function of IKKα in human physiology remains unknown since no biallelic variants disrupting its kinase activity have been reported. In this study, we present a homozygous germline missense variant in the kinase domain of IKKα, which is present in three children from two Turkish families. This variant, referred to as IKKαG167R, is in the activation segment of the kinase domain and affects the conserved (DF/LG) motif responsible for coordinating magnesium atoms for ATP binding. As a result, IKKαG167R abolishes the kinase activity of IKKα, leading to impaired activation of the non-canonical NF-κB pathway. Patients carrying IKKαG167R exhibit a range of immune system abnormalities, including the absence of secondary lymphoid organs, hypogammaglobulinemia and limited diversity of T and B cell receptors with evidence of autoreactivity. Overall, our findings indicate that, unlike a nonsense IKKα variant that results in early embryonic lethality in humans, the deficiency of IKKα's kinase activity is compatible with human life. However, it significantly disrupts the homeostasis of the immune system, underscoring the essential and non-redundant kinase function of IKKα in humans.
© 2024. The Author(s).
Conflict of interest statement
Figures
References
-
- Cildir, G., Low, K. C. & Tergaonkar, V. Noncanonical NF-kappaB Signaling in Health and Disease. Trends Mol. Med.22, 414–429 (2016). - PubMed
-
- Kinoshita, D. et al. Essential role of IkappaB kinase alpha in thymic organogenesis required for the establishment of self-tolerance. J. Immunol.176, 3995–4002 (2006). - PubMed
-
- Onder, L. et al. Alternative NF-kappaB signaling regulates mTEC differentiation from podoplanin-expressing precursors in the cortico-medullary junction. Eur. J. Immunol.45, 2218–2231 (2015). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- SRA/PRJNA1180092
Grants and funding
- 121S667/Türkiye Bilimsel ve Teknolojik Araştirma Kurumu (Scientific and Technological Research Council of Turkey)
- 2023-S-DTFA-003-QA25325/Hospital Research Foundation (The Hospital Research Foundation)
- 2023-S-DTFA-003-QA25325/Hospital Research Foundation (The Hospital Research Foundation)
- 2027602/Department of Health | National Health and Medical Research Council (NHMRC)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
