

Preclinical evaluation of a universal inactivated influenza B vaccine based on the mosaic hemagglutinin-approach
- PMID: 39551795
- PMCID: PMC11570629
- DOI: 10.1038/s41541-024-01014-8
Preclinical evaluation of a universal inactivated influenza B vaccine based on the mosaic hemagglutinin-approach
Abstract
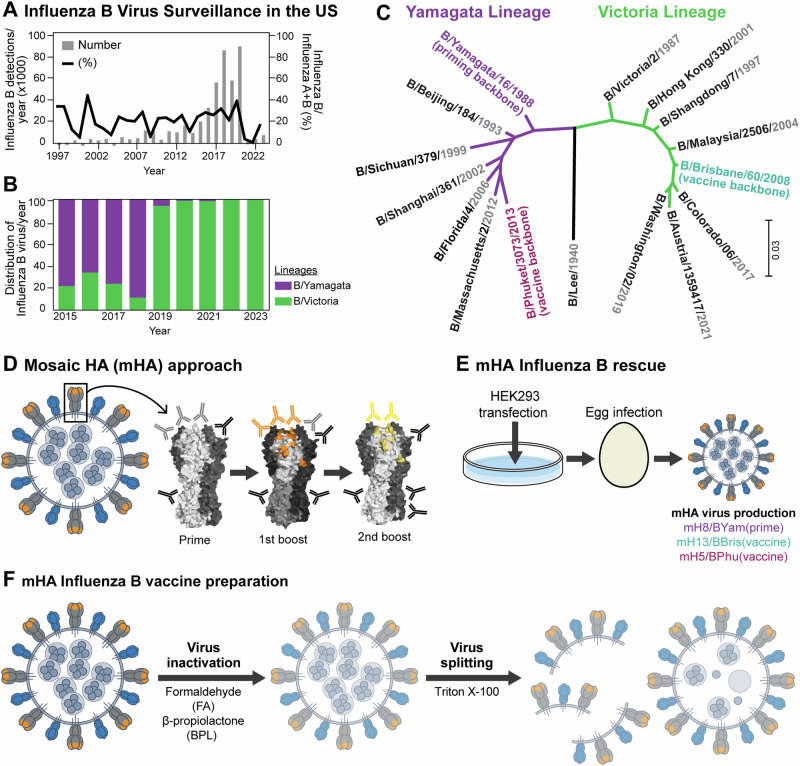
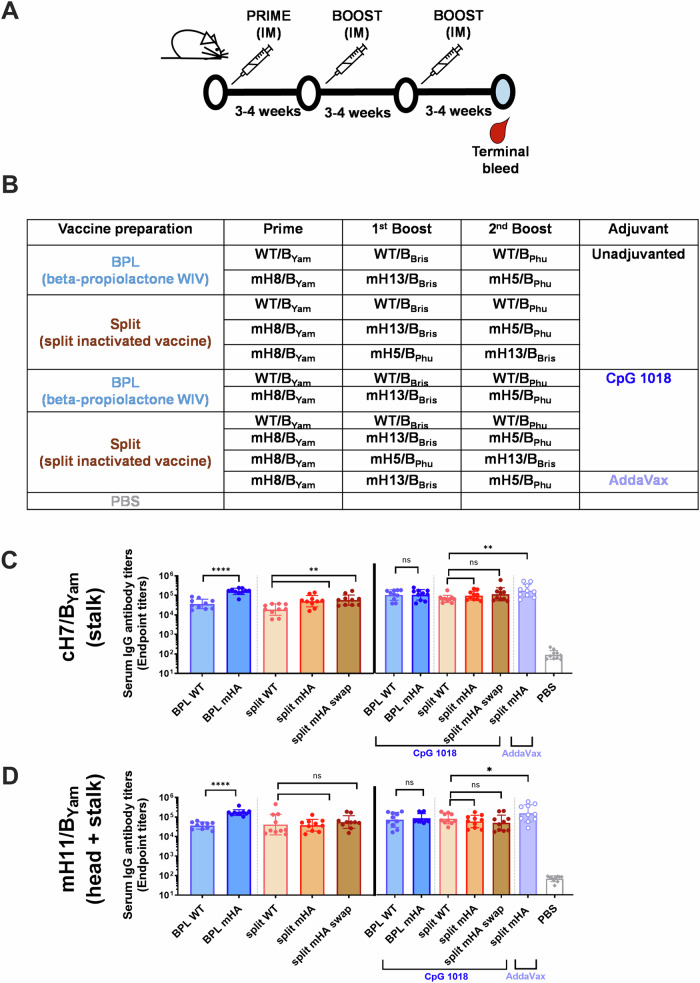
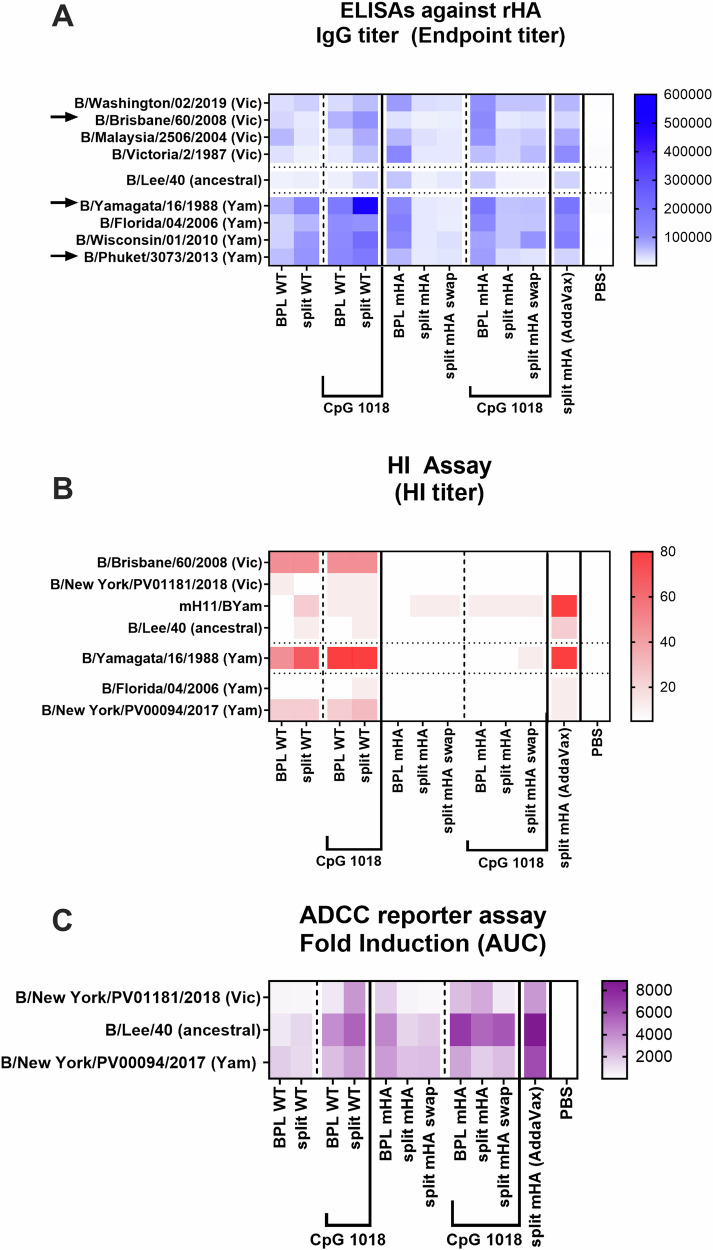
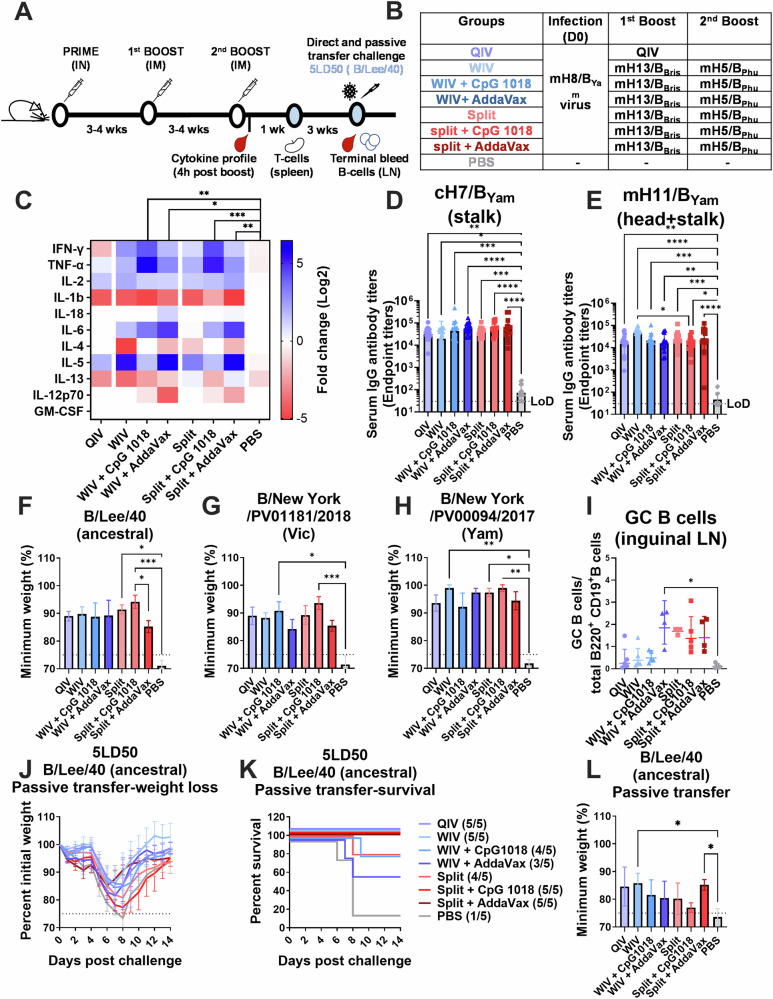
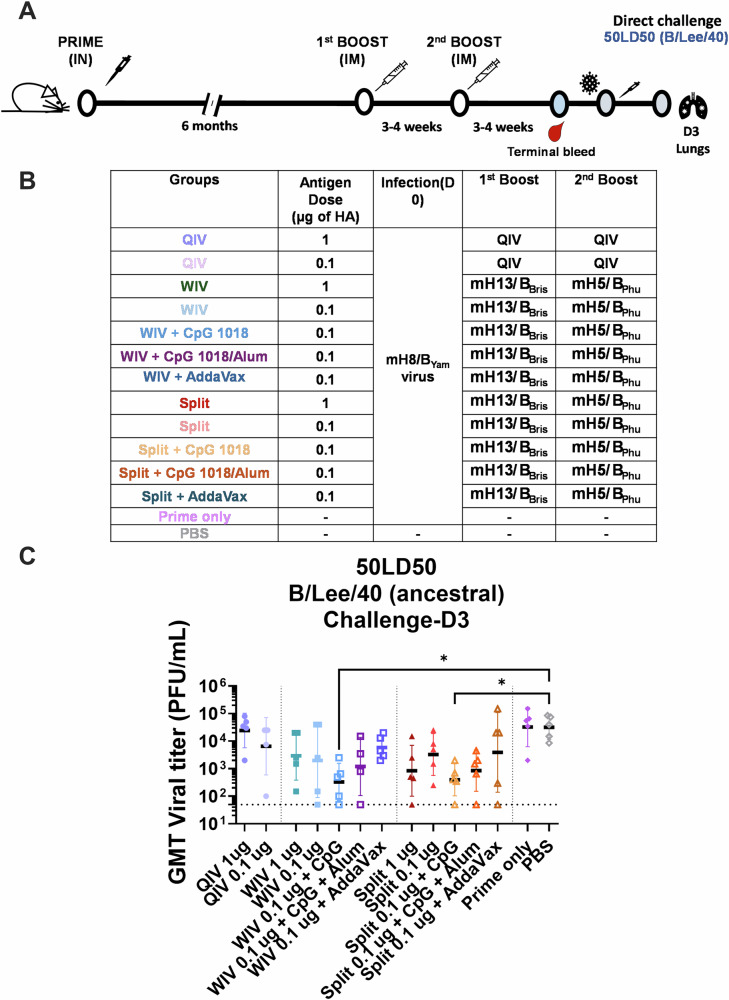
We have developed a new universal influenza B vaccination strategy based on inactivated influenza B viruses displaying mosaic hemagglutinins (mHAs). Recombinant mHA viruses were constructed by replacing the four major antigenic sites of influenza B virus HAs, with those from exotic avian influenza A virus HAs. Sequential vaccination of naïve mice with mHA-based vaccines elicited higher immune responses towards the immuno-subdominant conserved epitopes of the HA than vaccination with wildtype viruses. Among the different preparations tested, mHA split vaccines were less immunogenic than their whole inactivated virus counterparts. This lower immunogenicity was overcome by the combination with adjuvants. mHA split vaccines adjuvanted with a Toll-like receptor-9 agonist (CpG 1018) increased Th1 immunity and in vivo cross-protection, whereas adjuvanting with an MF59-like oil-in-water nano-emulsion (AddaVax) enhanced and broadened humoral immune responses and antibody-mediated cross-protection. The mHA vaccines with or without adjuvant were subsequently evaluated in mice that were previously immunized to closely mimic human pre-existing immunity to influenza B viruses and the contribution of innate and cellular immunity was evaluated in this model. We believe these preclinical studies using the mHA strategy represent a major step toward the evaluation of a universal influenza B virus vaccine in clinical trials.
© 2024. The Author(s).
Conflict of interest statement
Figures
References
-
- CDC. https://www.cdc.gov/flu/index.htm (2023).
Grants and funding
- R01 AI145870-03/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- CEIRR,75N93021C00014/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R01 AI145870/AI/NIAID NIH HHS/United States
- P01 AI097092/AI/NIAID NIH HHS/United States
- 75N93021C00014/AI/NIAID NIH HHS/United States
- CIVICs, 75N93019C00051/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- 75N93019C00051/AI/NIAID NIH HHS/United States
- P01 AI097092-07/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
LinkOut - more resources
Full Text Sources
