

This is a preprint.
Defective Neurogenesis in Lowe Syndrome is Caused by Mitochondria Loss and Cilia-related Sonic Hedgehog Defects
- PMID: 39553960
- PMCID: PMC11565974
- DOI: 10.1101/2024.11.01.621496
Defective Neurogenesis in Lowe Syndrome is Caused by Mitochondria Loss and Cilia-related Sonic Hedgehog Defects
Abstract
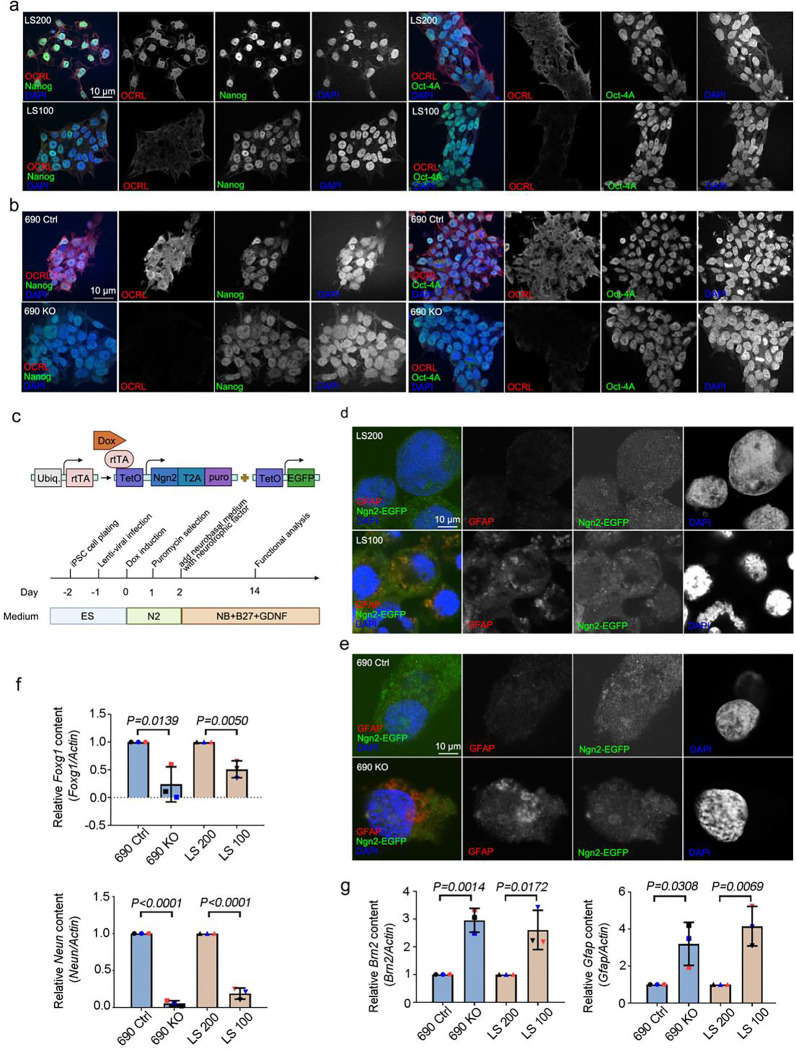
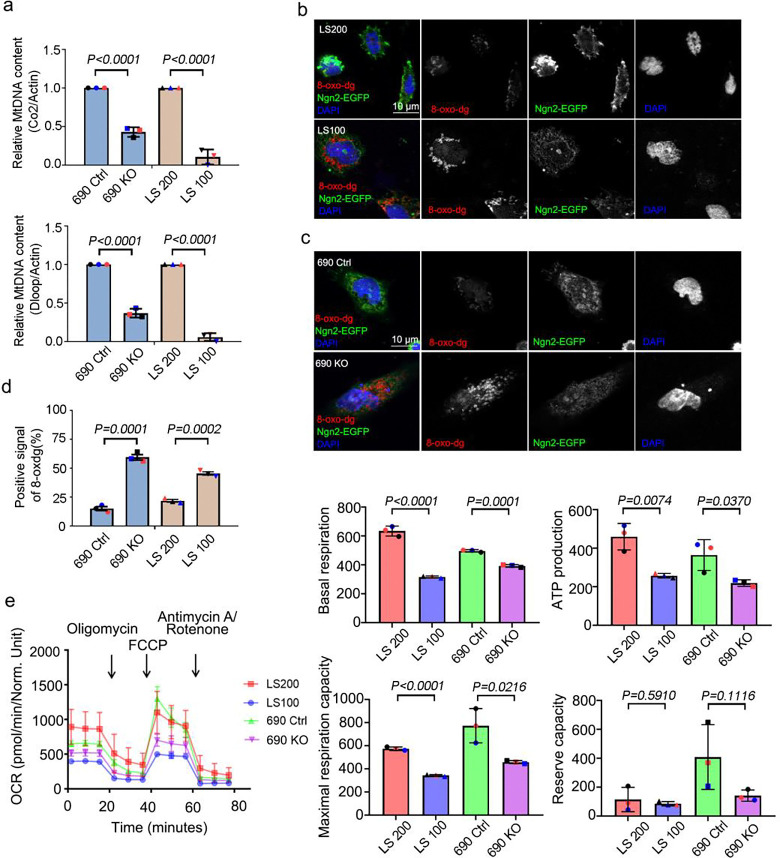
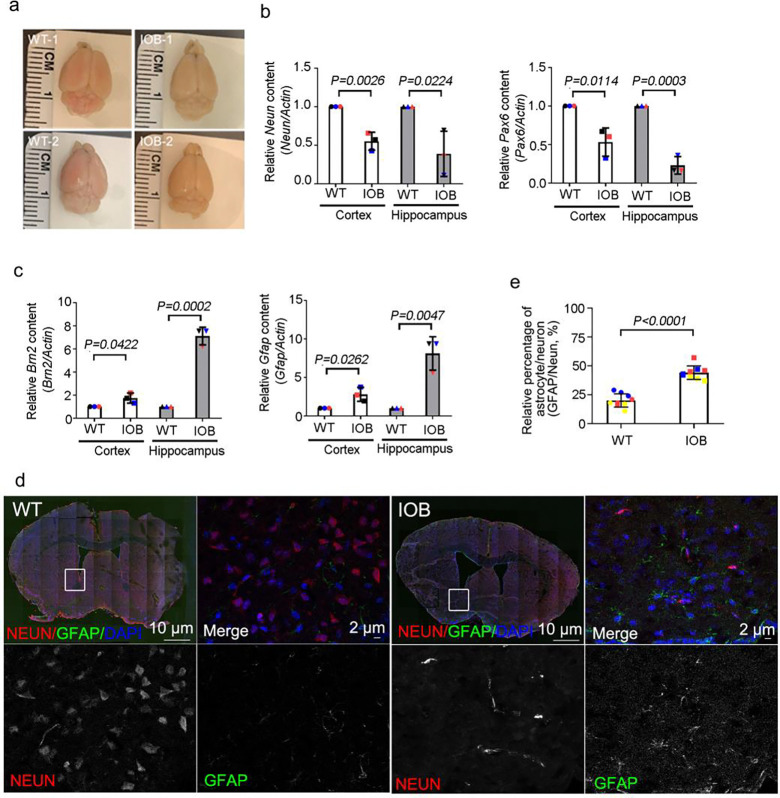
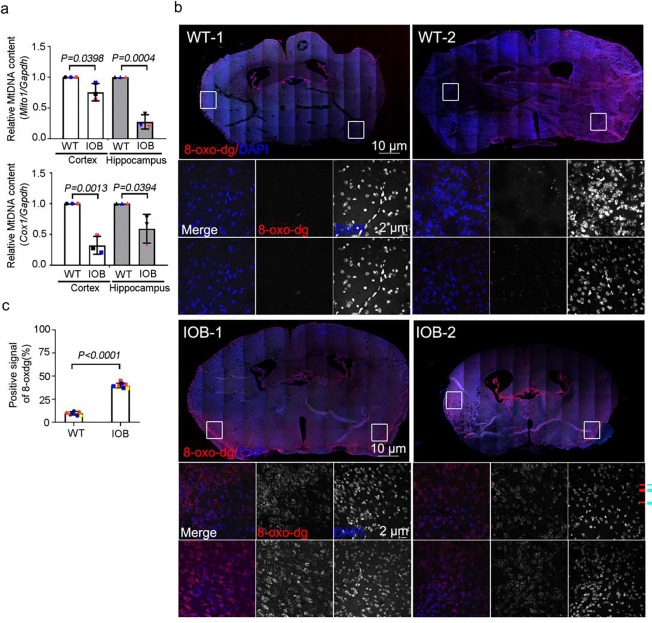
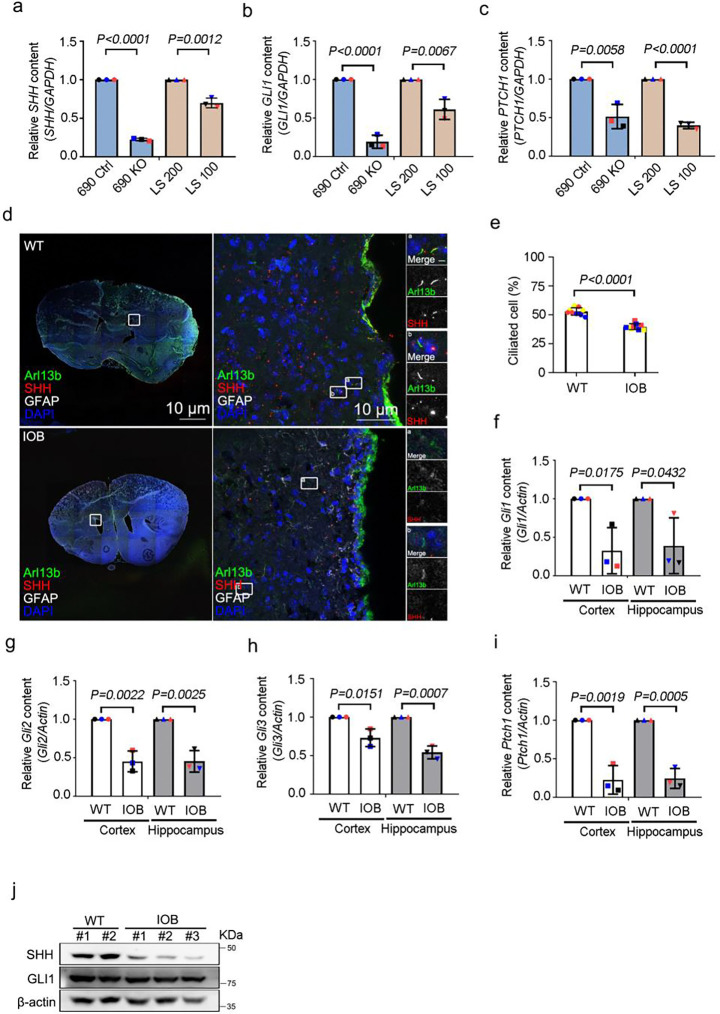
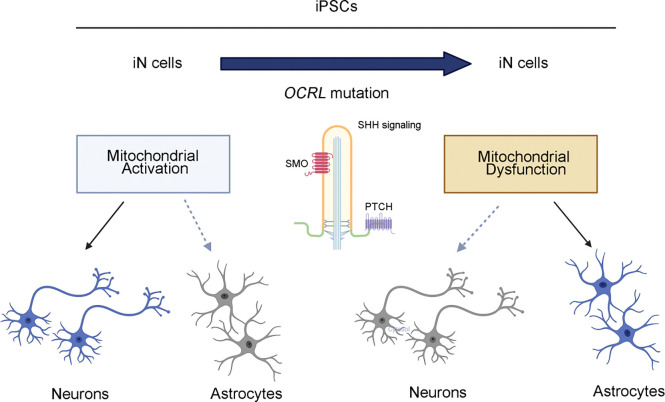
Human brain development is a complex process that requires intricate coordination of multiple cellular and developmental events. Dysfunction of lipid metabolism can lead to neurodevelopmental disorders. Lowe syndrome (LS) is a recessive X-linked disorder associated with proximal tubular renal disease, congenital cataracts and glaucoma, and central nervous system developmental delays. Mutations in OCRL, which encodes an inositol polyphosphate 5-phosphatase, lead to the development of LS. The cellular mechanism responsible for neuronal dysfunction in LS is unknown. Here we show depletion of mitochondrial DNA and decrease in mitochondrial activities result in neuronal differentiation defects. Increased astrocytes, which are secondary responders to neurodegeneration, are observed in neuronal (iN) cells differentiated from Lowe patient-derived iPSCs and an LS mouse model. Inactivation of cilia-related sonic hedgehog signaling, which organizes the pattern of cellular neuronal differentiation, is observed in an OCRL knockout, iN cells differentiated from Lowe patient-derived iPSCs, and an LS mouse model. Taken together, our findings indicate that mitochondrial dysfunction and impairment of the ciliary sonic hedgehog signaling pathway represent a novel pathogenic mechanism underlying the disrupted neuronal differentiation observed in LS.
Keywords: Cilia formation; Lowe Syndrome; Mitochondria; Neuronal differentiation; OCRL.
Conflict of interest statement
Conflict of Interest Statement The authors declare no competing interests.
Figures
References
-
- Murdock N. and Chou E., Oculocerebrorenal Syndrome, in StatPearls. 2024: Treasure Island (FL) ineligible companies. Disclosure: Eva Chou declares no relevant financial relationships with ineligible companies.
-
- Sharma S., Skowronek A., and Erdmann K.S., The role of the Lowe syndrome protein OCRL in the endocytic pathway. Biol Chem, 2015. 396(12): p. 1293–300. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources